拟南芥RPS14基因的克隆表达及功能初步研究
2013-12-23范艳荣刘莹张飞云
范艳荣 刘莹 张飞云
(首都师范大学生命科学学院,北京 100048)
核糖体是合成蛋白质的重要细胞器,包括40S小亚基和60S大亚基。这些亚基分别有4种不同类型的RNA和80种结构不同的蛋白质组成。RPS14(Ribosomal protein S14)是40S亚基的组成成分,这个蛋白属于S11P核糖体蛋白家族[1]。根据已有研究,人类、酵母等真核生物体内的RPS14基因主要负责RNA转录后加工,如SSU-rRNA(small subunit rRNA)的加工成熟与组装,从而调控蛋白质的翻译过程。在中国仓鼠的卵巢细胞中,该基因的突变能导致产生蛋白合成抑制剂,抵抗催吐剂的合成。
拟南芥腺苷酸激酶6(Arabidopsis thaliana Adenylate kinase 6,AAK6)是一种腺苷酸单磷酸激酶,广泛存在于生物体中。AK6基因已在大多数动物,如人类、果蝇、线虫以及酵母中有了较深入的研究,近年来人们开始探究AK6基因在植物体内的特性与功能。以模式植物拟南芥为研究对象,已证实AK6蛋白具有腺苷酸激酶活性,能可逆地催化γ磷酸基团的转移(通常作用于ATP),使ATP和AMP反应生成两分子的ADP,以此来维持细胞内的能量平衡[2-4]。同时,脊椎动物、果蝇、酵母菌,以及植物细胞核中存在一种亚核结构柯浩体,它能够行使mRNA和rRNA前体的拼接和剪接功能。在对人类AK6基因(HAK6/CBRF)的初步研究表明,AK6基因定位于柯浩体上,故又称为柯浩体调节因子,其很可能在柯浩体上发挥重要作用,与真核细胞中核糖体的组装相关[5,6]。通过前期的研究,从aak6-/aak6-突变体植株与野生型植株的生长状况对比可以看出,突变体明显矮小[6,7]。
RPS14基因参与RNA的转录后加工过程,推测其可能影响植株的生长状况。本研究拟用体外pulldown技术,并结合SDS-PAGE和Western blot检测,旨在探究拟南芥中RPS14蛋白与AK6蛋白是否存在相互作用,找到植株矮化的内在线索。
1 材料与方法
1.1 材料
1.1.1 菌株和质粒 pEASY-T3、pGEX-6P-1载体、pET-28a载体、大肠杆菌(Escherichia coli)DH5α与TOP10、BL21(DE3)均为本实验室保存。
1.1.2 试剂 Trizol试剂购自美国TEL-TEST Inc。常用化学无机盐等试剂均为北化分析纯试剂。逆转录试剂盒购自Fermerters公司。高保真PCR试剂盒、随机引物购自上海生工生物工程公司。T-A cloning 试剂盒购自全式金公司。各种构建载体所需工具酶(如限制性内切酶、T4连接酶等)购于TaKaRa公司。质粒小提试剂盒、琼脂糖凝胶DNA回收试剂盒购自QIN-GEN公司。
1.2 方法
1.2.1 RNA提取与检测及反转录 以拟南芥的叶片为材料,用Trizol法提取RNA,并用反转录试剂盒反转录成cDNA。
1.2.2 RPS14基因的扩增 根据GenBank中2g36160序列利用PrimerPremier5.0设计引物,上游EcoR I 酶切位点,下游加入Xho I 酶切位点,引物序列为:
上游引物:5'-GAATTCATGTCAAAGAGAAAGACTAAAGAGC-3';下游引物:5'-CTCGAGTCAGAGCCTTCTTCCTCTTCTAC-3'。
以制备好的拟南芥cDNA为模板,扩增体系为20 μL反应程序:94℃预变性5 min;94℃变性30 s,58℃退火40 s,72℃延伸1 min,30个循环;72℃ 5 min。反应结束后,取出5 μL进行电泳,检测是否扩增出目的片段,检测到目的片段后用胶回收试剂盒进行回收。
1.2.3 原核表达质粒的构建和鉴定 将纯化后的PCR产物和pEASY-T3连接,用限制性内切酶 EcoR I和Xho I进行双酶切,胶纯化以后将PCR产物与pGEX-6P-1线性片段连接,16℃连接过夜,连接体系如表1所示。
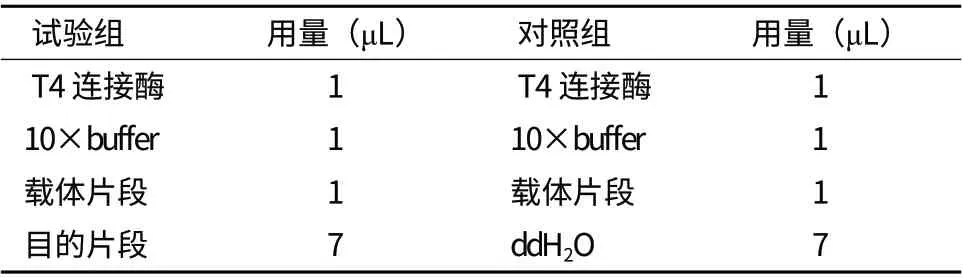
表1 PCR产物与pGEX-6P-1线性片段连接体系
将连接产物转化至大肠杆菌BL21(DE3)感受态细胞中,37℃活化1.5 h后涂LB Amp抗性平板,置37℃倒置培养。转化后,用抗生素筛选后的单克隆进行如下程序检测:菌落PCR筛选—测序,最终找出正确的重组克隆。
1.2.4 重组蛋白在大肠杆菌中的表达和纯化 将测序结果正确的重组质粒分别转化BL21(DE3)感受态细胞表达菌株,用0.5 mmol/L的IPTG诱导,16℃ 23 h。5 000 r/min,30 min离心,弃上清。用5 mL PBS悬浮沉淀。然后超声破碎,4℃ 12 000 r/min 离心30 min,收集上清,上清和沉淀分别留少许进行SDS-PAGE检测。在上清中加入适量GST beads,4℃旋转令其吸附蛋白2.5 h;2 000 r/min,3 min离心弃上清;加入至少50 mL PBS,轻摇至beads悬浮于溶液中,2 000 r/min,3 min离心,弃上清;重复用PBS洗beads两次; 加入1 mL GST Elution Buffer,轻摇10 min;2 000 r/min,3 min离心,收集上清;重复用GST Elution Buffer洗两次,收集上清进行SDSPAGE检测。通过SDS-PAGE电泳检测。
1.2.5 过表达的拟南芥AK6蛋白的获得 AK6引物:上游引物:5'-GGAATTCCATATGGCGAGACGTAACCGTGGG-3'(Nde I);下游引物:5'-CGGGATCCTCAGGGTTGCCATGCATTA-3'(BamH I)。其 他 方 法 同RRS14蛋白的制备。
1.2.6 体外pull-down寻找与RPS14相互作用的蛋白及免疫检测 用50 μL GST beads和1 mL GST-RPS14蛋白在4℃孵育,将过表达的HIS-AK6蛋白混合液加入经处理的GST beads中,在4℃旋转混合孵育2 h,用冰冷的细胞裂解液洗涤重复洗3次,SDS-PAGE检测。取胶,在冰浴中转PVDF膜2 h,将膜从电转槽中取出,用去离子水与PBST稍加漂洗,浸没于封闭液中缓慢摇荡1 h。加含HIS-Tag标签的一抗,4℃摇动孵育2 h。用PBST洗膜3次,每次5-10 min。加入相应的二抗后,用PBST再洗膜3次,每次5-10 min。最后进行自显影。
2 结果
2.1 总RNA的提取及反转录
用Trizol从拟南芥的叶片中提取RNA,根据检测的浓度利用反转录试剂盒将RNA反转录成cDNA,保存在-20℃备用,结果如图1所示。

利用特异的上下游引物通过PCR程序从拟南芥cDNA中扩增目的条带,1%的琼脂糖凝胶电泳检测扩增出的PCR产物,结果(图2)表明获得了与预期片段大小相符的清晰的目的条带。
2.2 原核重组表达质粒的构建和鉴定
将切胶回收的目的基因PCR产物与pEASYT3载体连接,用限制性内切酶EcoR I和Xho I 37℃双酶切pEASY-T3-RPS14和pGEX-6P-1,结果(图3)显示连接成功。把切下的目的基因和双酶切后的pGEX-6P-1载体16℃过夜连接,转化,挑取健壮的单克隆摇菌,用菌液PCR检测出单一条带,结果(图4)显示为阳性菌株。将阳性菌株菌液送去测序。测序结果与NCBI的相关序列比对完全正确。


2.3 原核重组表达载体的表达、可溶性分析及蛋白纯化
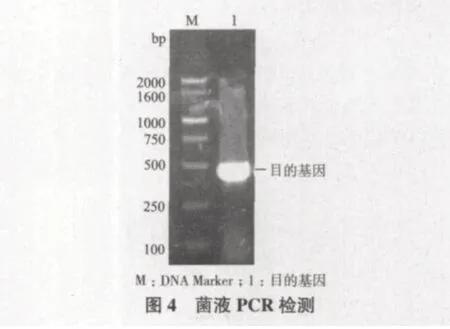
将测序正确的重组质粒分别转化到BL21(DE3)中,分别挑取健壮的单克隆培养至OD600=0.6,加入诱导剂IPTG在16℃诱导23 h;5 000 r/min,30 min离心,弃上清。用5 mL PBS悬浮沉淀。然后超声破碎,上清和沉淀用SDS-PAGE检测,如图5-A所示。蛋白纯化后,经聚丙烯酰胺凝胶电泳检测得到比较单一的条带,结果如图5-B所示。
2.4 过表达拟南芥AK6蛋白
获得AK6蛋白需构建原核表达质粒pET-28a-AK6,其中pET-28a中带有HIS标签,便于后续试验中进行Western blot以鉴定AK6蛋白的存在,从而证明拟南芥中RPS14蛋白与AK6蛋白发生相互作用。
2.5 核糖体组成蛋白RPS14可能与AK6相互作用
将纯化后的HIS-AK6和GST-RPS14融合蛋白与GST beads共同孵育,经离心收集洗脱复合物和洗涤后,再加入过量GSH获得相互作用蛋白的复合物,去上清用SDS-PAGE检测(图6)。取胶,在冰浴中转PVDF膜2 h,最后进行自显影。
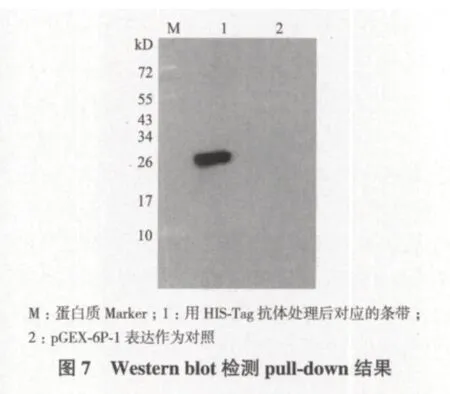
Western blot检测结果(图7)显示,凝胶泳道1转移膜上出现明显目的条带,且条带位于约26 kD处;而对照组凝胶泳道2转移膜上无目的条带出现。由于试验组pull-down收集的上清液蛋白中,只有AK6融合蛋白带有HIS标签,免疫检测加入识别HIS标签的抗体,使HIS-AK6融合蛋白与之结合,最后显色位置进一步证明上清液蛋白中确实存在AK6蛋白,并确定SDS-PAGE结果中出现的26 kD处条带是AK6蛋白,从而也进一步证明拟南芥中RPS14蛋白与AK6蛋白确实存在相互作用,共同调控细胞能量代谢平衡,这在AK6缺失导致拟南芥生长矮化的机制中可能发挥了重要作用。

3 讨论
拟南芥腺苷酸激酶6(AA6)是P-loop激酶家族成员,该激酶家族具有典型、保守的Walk A和Walk B结构域,其中Walk A结构域能够结合NTP分子,Walk B结构域能够结合重要的辅因子Mg2+,能使NTP+NMP=2NDP。核苷酸的平衡对多种细胞内的功能非常重要,如细胞内能量的代谢及DNA和RNA的合成等。
由于AK6是腺苷酸激酶,在体外试验中,除酵母外,人类、线虫、果蝇AK6的同源蛋白都具有激酶活性[8,9],能使所有类型的NTPs和dNTPs充当磷酸供体,最适磷酸供体是ATP和dATP。同时,与拟南芥野生型植株相比,aak6-/aak6-纯合突变体植株表现出明显的矮化[10],暗示AK6基因的缺失影响了细胞的形态和生长。
研究发现,酵母RPS14蛋白C端精氨酸突变成丙氨酸时,与Fap7(AK6的同源蛋白)缺失的表型相似,证实YAK6(Fap7)能与核糖体蛋白质RPS14互作,共同促进20S rRNA前体中D位点的剪接过程,使20S rRNA前体剪接为成熟的18S rRNA,再进一步组装为40S核糖体小亚基。如果把Fap7上Walk A和Walk B 结构域中几个保守的氨基酸突变以后,能明显抑制20S rRNA前体的剪接和细胞的生长[11],说明RPS14基因对RNA的编辑起重要作用。
推测在拟南芥中,RPS14和AK6也可能存在相互作用。将GST空载,GST-RPS14融合蛋白和HISAK6融合蛋白在细菌中过表达,并亲和层析纯化。将纯化后的GST空载,GST-RPS14蛋白分别与HISAK6蛋白在冰上同GST beads孵育,纯化的GST空载作为负对照。用SDS-PAGE检测,结果显示GSTRPS14和HIS-AK6特异性的结合,而负对照组不发生结合。Western blot进一步证实GST-RPS14和HISAK6在体外有相互作用。
4 结论
本研究选用模式植物拟南芥作为试验材料克隆了RPS14基因和AK6基因,并获得其可溶性蛋白GST-RPS14和HIS-AK6。运用体外GST pull-down技术得出,拟南芥 RPS14蛋白与AK6蛋白存在相互作用。
[1] Granneman S, Nandineni MR, Baserga SJ. The Putative NTPase Fap7 mediates cytoplasmic 20S pre-rRNA processing through a direct interaction with RpS14[J]. Mol Cell Biol, 2005, 25(23):10352-10364.
[2] Whitford PC, Miyashita O, Levy Y, Onuchic JN. Conformational transitions of adenylate kinase:Switching by cracking[J]. J Mol Biol, 2007, 366(5):1661-1671.
[3] Van Rompay AR, Johansson M, Karlsson A. Identifucation of a novel human adenylate kinase. cDNA cloning, expression analysis, chromosome localization and characterization of the recombinant protein[J]. Eur J Biochem, 1999, 261:509-517.
[4] Zancani M, CasoloV, VianelloA, Macri F. Involvement of apyrase in the regulation of the adenylate pool by adenylate kinase in plant mitochondria[J]. Plant Sci, 2001, 161(5):927-933.
[5] Zhai R, Meng G, Zhao Y, et al. A novel nuclear-localized protein with special adenylate kinase properties from Caenorhabditis elegans[J]. FEBS Lett, 2006, 580(16):3811-3817.
[6] Ren H, Wang L, Bennett M, et al. The crystal structure of human adenylate kinase 6:An adenylate kinase localized to the cell nucleus[J]. Proc Natl Acad Sci, 2005, 102(2):303-308.
[7] Lange PR, Geserick C, Tischendorf G, Zrenner R. Functions of chloroplastic adenylate kinases in Arabidopsis[J]. Plant Physiology, 2007, 146(2):492-504.
[8] Juhnke H, Charizanis C, Latifi F, et al. The essential protein fap7 is involved in the oxidative stress response of Saccharomyces cerevisiae[J]. Mol Microbiol, 2000, 35(4):936-948.
[9] Carrari F, Coll-Garcia D, Schauer N, et al. Deficiency of a plastidial adenylate kinase in Arabidopsis results in elevated photosynthetic amino acid biosynthesis and enhanced growth[J]. Plant Physiologl, 2005, 137(1):70-82.
[10] Zhang J, Zhang F, Zheng X. Depletion of hCINAP by RNA interference causes defects in Cajal body formation, histone transcription, and cell viability[J]. Cellular and Molecular Life Sciences, 2010, 67(11):1907-1918.
[11] Meng G, Zhai R, Chen B, et al. Identification and characterization of human adenylate kinase 6 homologues from Drosophila melanogaster[J]. Biochemistry(Moscow), 2008, 73(1):38-43.
