硫氧还蛋白硝基化在多柔比星诱导的乳鼠心肌细胞凋亡中的作用
2013-12-23李悦山
王 斌, 李悦山
(广州医科大学药理教研室,广东 广州510182)
多柔比星(doxorubicin,DOX)属蒽醌类抗生素,是临床上一种高效、广谱抗肿瘤药物,但其严重的心脏毒性限制了它的应用[1]。多柔比星心脏毒性的一个严重表现是诱导心肌细胞凋亡的发生[2]。研究发现,细胞内蛋白质硝基化修饰可以直接影响其生物学功能[3],而多柔比星可以提高细胞内的蛋白质硝基化水平[4]。细胞内蛋白质的硝基化修饰是否是多柔比星诱导细胞凋亡的因素及其作用机制,目前尚无明确的报道。
硫氧还蛋白(thioredoxin,Trx)是生物体内广泛存在的一种多功能小分子蛋白,因其在各个器官普遍存在且发挥重要作用而备受研究者的关注。临床和动物研究发现Trx 活性与细胞凋亡密切相关。研究已经证实,硝基化修饰的Trx 活性降低,导致其抑制凋亡信号调节激酶1 (apoptosis signal-regulating kinase 1,ASK1)活性的能力降低[5-7],进而激活其下游信号通路。本实验我们以多柔比星刺激心肌细胞模型,观察心肌细胞Trx 硝基化水平的变化以及其对Trx-ASK1 等凋亡相关信号分子的影响,为揭示多柔比星心脏毒性和细胞凋亡的发生机制提供依据。
材 料 和 方 法
1 材料
1.1 主要试剂 多柔比星和细胞级二甲基亚砜(dimethyl sulfoxide,DMSO)购于Sigma;锰(III)四(1-甲基-4-吡啶基)卟啉[Mn(III)tetrakis(1-methyl-4-pyridyl) porphyrin,MnTMPyP]购于 Alexis;高 糖DMEM 培养基、胎牛血清和胰蛋白酶购于Gibco;MTT 细胞增殖及细胞毒性检测试剂盒、caspase-3 分光光度法检测试剂盒和细胞凋亡荧光Hoechst 33258试剂盒购于南京凯基生物;蛋白定量试剂盒和发光液购于Pierce;Trx 和硝基酪氨酸(nitrotyrosine,NT)单克隆抗体以及ASK1 多克隆抗体购于Santa Cruz;聚ADP 核糖聚合酶1 剪切片段[cleaved poly(ADPribose)polymerase-1,PARP-1]、磷酸化ASK1 (p-ASK1)、p38 丝裂原活化蛋白激酶(p38 mitogen-activated protein kinase,p38 MAPK)和磷酸化p38 MAPK(p-p38 MAPK)抗体购于Cell Signaling Technology。
1.2 动物 SPF 级Sprague-Dawley 新生大鼠(1 ~3 d,雌雄不限)由广州中医药大学动物中心提供。
2 方法
2.1 心肌细胞分离培养 乳鼠心肌细胞原代培养方法参照Simpson 等[8]的方法改进而得。新生SD乳鼠用1%新洁尔灭洗净后,在无菌超净台中打开胸腔,迅速取出左室心肌组织浸入冰冷的D-Hanks 液中,洗净心室内血液,剪成(0.5 ~1)mm ×1 mm ×1 mm 大小的组织块,冲洗数次;将剪碎的心肌组织转移至锥形瓶中,加0.125%胰蛋白酶液3 ~4 mL,37℃恒温磁力搅拌50 r/min,消化8 ~10 次,每次6 ~8 min。从第3 次消化起,收集上清(每次消化后,用吸管轻轻吹打30 s,静置约2 min,用含30%新生牛血清的培养基终止消化,收集细胞)。1 500 r/min 离心10 min,弃上清。收集沉淀加入到含15% 胎牛血清的DMEM 培养基中重悬细胞。将细胞悬液培养于培养皿中,于37 ℃、5% CO2水套式培养箱中差速贴壁2 h 后,小心吸出细胞悬浮液并计数。根据需要稀释到一定密度(5 ×105~1 ×106/L)接种于培养皿(或孔板)中,24 h 后换为含15%胎牛血清加胰岛素(10 mg/L)、转铁蛋白(10 mg/L)、维生素B12(1.5 μmol/L)和BrdU(0.1 mmol/L)的DMEM 培养基。96 h 后换用不含BrdU 的上述无血清培养基。倒置相差显微镜下观察细胞生长的情况。细胞形成放射状排列的细胞簇,呈同步性搏动,搏动细胞大于95%,则可进行以下实验。
2.2 心肌细胞活力测定 参照南京凯基生物的MTT 细胞增殖及细胞毒性检测试剂盒说明书,通过酶标仪550 nm 波长检测吸光度值,计算心肌细胞的存活率。
2.3 细胞浆中caspase-3 活性的测定 参照南京凯基生物的caspase-3 分光光度法检测试剂盒说明书,通过酶标仪405nm 波长检测吸光度值,检测caspase-3 的活化程度。
2.4 心肌细胞凋亡荧光显微镜观察 参照南京凯基生物的细胞凋亡荧光Hoechst 33258 试剂盒说明书,荧光显微镜下,检测细胞凋亡情况。
2.5 检测Trx-ASK1 的分离和Trx-NT 的含量 用多克隆ASK1 抗体免疫沉淀(immunoprecipitation,IP)法获取细胞中的总ASK1,Trx 单克隆抗体Western blotting 检测Trx-ASK1;用Trx 单克隆抗体IP 法获取细胞内总Trx,NT 多克隆抗体Western blotting 检测Trx-NT,具体方法参照Pierce 公司的IP 试剂盒说明书。
2.6 检测cleaved PARP-1、p-ASK1( Ser967) 、p38 MAPK 和p-p38 MAPK 蛋白水平 提取细胞总蛋白后,BCA 法测定蛋白浓度,Bio-Rad 垂直电泳,转膜,5%脱脂奶粉封闭1 h,4 ℃冰箱孵育1 抗(1∶1 000),过夜,TBST 洗膜3 次,每次8 min,加Ⅱ抗(1∶1 000)室温孵育2 h,压片法曝光成像。结果用ImageJ 图像分析系统进行分析。
3 统计学处理
用SPSS 16.0 统计软件分析,数据用均数±标准差(mean ±SD)表示,组间比较采用单因素方差分析(ANOVA),组间两两比较采用最小显著性差异法(LSD 法),以P <0.05 为差异有统计学意义。
结 果
1 不同浓度多柔比星作用对细胞活力的影响
给予0、0.1、0.3、1、3 和10 μmol/L 多柔比星作用心肌细胞24 h 后,MTT 方法检测细胞活力。1、3与10 μmol/L 多柔比星组MTT 值下降,与对照组比较均有显著差异(P <0.05),见图1A。
2 胞浆caspase-3 活性的检测
不同浓度多柔比星作用心肌细胞24 h 后,检测胞浆caspase-3 活性。与对照组相比,1、3 和10 μmol/L 多柔比星组的caspase-3 活化度升高,分别为(171.53 ±18.25)%、(176.03 ±10.31)%和(187.00±18.39)%(P <0.05),见图1B。
3 Cleaved PARP-1 蛋白表达的检测
多柔比星作用心肌细胞24 h 后,Western blotting检测细胞内cleaved PARP-1 蛋白的表达。与对照组相比,1、3 和10 μmol/L 多柔比星组的cleaved PARP-1 表达明显增加(P <0.05 或P <0.01),见图1C。

Figure 1. Effects of different concentrations of doxorubicin (DOX)on the viability and apoptosis of neonatal rat cardiomyocytes. A:cell viability;B:activity of caspase-3;C:expression of cleaved PARP-1 protein detected by Western blotting.Mean±SD.n=8 in A;n=3 in B and C. * P <0.05,**P <0.01 vs 0 μmol/L.图1 不同浓度DOX 对心肌细胞活力和凋亡相关指标的影响
4 Trx 硝基化的检测
多柔比星作用心肌细胞24 h 后,检测细胞内Trx-NT 的表达。与对照组相比,1、3 和10 μmol/L 多柔比星组的Trx-NT 表达明显增加(P <0.05),见图2。
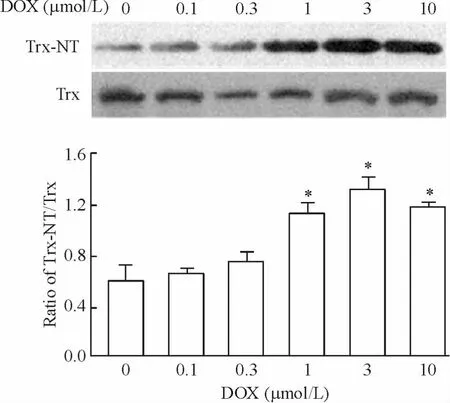
Figure 2. Effects of different concentrations of DOX on the nitration of thioredoxin (Trx). NT:nitrotyrosine. Mean ±SD.n=3. * P <0.05 vs 0 μmol/L.图2 不同浓度DOX 对Trx 硝基化的影响
5 不同浓度MnTMPyP 作用对细胞活力的影响
预孵育不同浓度MnTMPyP 2 h,再加1 μmol/L多柔比星,24 h 后检测细胞活力。结果显示,1 μmol/L 多柔比星组较对照组MTT 值下降(P <0.05),30 和100 μmol/L 的MnTMPyP 可以显著抑制多柔比星对细胞的损伤,与多柔比星组MTT 值比较有显著差异(P <0.05),见图3A。
6 细胞凋亡荧光显微镜观察
预孵育30 μmol/L MnTMPyP 2 h,再给予1 μmol/L 多柔比星,24 h 后进行Hoechst 33258 荧光染色。对照组细胞呈低蓝色,多柔比星组细胞呈高蓝色,亮度明显增加,可观察到明显的细胞核固缩,具有凋亡的显著特征;与多柔比星组相比,MnTMPyP组凋亡有所改善,见图3B。
7 MnTMPyP 作用后caspase-3 活性的检测
预孵育30 μmol/L MnTMPyP 2 h,再给予1 μmol/L 多柔比星作用心肌细胞24 h 后,检测胞浆中caspase-3 的活性。相比于对照组,1 μmol/L 多柔比星组的caspase-3 活化度为(175.22±16.22)%(P <0.05);与多柔比星组相比,MnTMPyP 可以显著抑制caspase-3的活化[126.30±9.62)%,P <0.05],见图3C。
8 MnTMPyP 作用后检测cleaved PARP-1 的表达
预孵育30 μmol/L MnTMPyP 2 h,再给予1 μmol/L 多柔比星24 h 后,与对照组相比,多柔比星组cleaved PARP-1 表达明显增加(P <0.01);与多柔比星组相比,MnTMPyP 可以明显降低cleaved PARP-1 表达(P <0.05),见图3D。

Figure 3. Effects of MnTMPyP (MnT)on the viability and apoptosis of neonatal rat cardiomyocytes induced by DOX. A:cell viability;B:Hochest 33258 staining;C:activity of caspase-3;D:expression of cleaved PARP-1 detected by Western blotting.Mean±SD.n=8 in A;n=3 in B ~D. #P <0.05,##P <0.01 vs control;* P <0.05 vs DOX.图3 MnTMPyP 对多柔比星诱导的心肌细胞活力和凋亡的影响
9 免疫沉淀法检测Trx 硝基化程度
多柔比星作用24 h 后,与正常组比较,多柔比星组Trx 硝基化程度显著增加(P <0.05);与多柔比星组相比,MnTMPyP+多柔比星组硝基化程度显著降低(P <0.05),见图4A。
10 免疫沉淀检测Trx 与ASK1 的结合程度
预孵育30 μmol/L MnTMPyP 2 h,再给予1 μmol/L 多柔比星作用24 h 后,多柔比星组与正常组比较Trx-ASK1 分离增多(P <0.05);与多柔比星组相比,MnTMPyP +多柔比星组Trx-ASK1 分离减少,见图4B。

Figure 4. Effects of MnTMPyP(MnT)on nitration of Trx(A)and dissociation of Trx-ASK1(B)induced by DOX. NT:nitrotyrosine.Mean±SD.n=3. #P <0.05 vs control;* P <0.05 vs DOX.图4 MnTMPyP 对多柔比星诱导的Trx 硝基化和Trx-ASK1 分离的影响
11 Western blotting 检测细胞p-ASK1 (Ser967)
心肌细胞给予1 μmol/L 多柔比星作用24 h 后,与正常组比较,p-ASK1(Ser967)的表达显著减少(P <0.05);与多柔比星组相比,MnTMPyP +多柔比星组p-ASK1(Ser967)表达显著增加(P <0.05),见图5A。
12 Western blotting 检测细胞p38 MAPK 的活化
心肌细胞给予1 μmol/L 多柔比星作用30 min后,与正常组比较,p-p38 MAPK 表达显著增加(P <0.05);与多柔比星组相比,MnTMPyP +多柔比星组p38 MAPK 磷酸化程度显著降低(P <0.05);各组之间p38 MAPK 的表达无显著差异,见图5B。

Figure 5. Effects of MnTMPyP (MnT)on the activation of ASK1 (A)and p38 MAPK (B)induced by DOX. Mean ±SD. n =3.#P <0.05 vs control;* P <0.05 vs DOX.图5 MnTMPyP 对多柔比星诱导的ASK1 和p38 MAPK 活化的影响
讨 论
本实验应用多柔比星体外诱导乳鼠心肌细胞凋亡模型,观察和探讨此过程Trx 的硝基化情况以及凋亡相关信号分子的变化,为多柔比星导致的心肌损伤和细胞凋亡的具体机制提供依据。
多柔比星是一种蒽醌类抗生素,在临床上是一种有效的广谱抗肿瘤药物,但是在临床应用过程发现有很强的心脏毒性,可以导致各种心肌病甚至心力衰竭的发生。研究发现多柔比星导致心肌细胞凋亡是心肌细胞流失和心功能失常很重要的一方面[9]。目前已有众多关于多柔比星诱导心肌细胞凋亡机制的研究报道。线粒体功能紊乱,细胞内凋亡调节相关蛋白如P53 和Bcl-2 家族功能异常,血红素加氧酶下调,细胞内caspases 募集反应凋亡抑制蛋白的表达降低等[9],都与多柔比星的细胞毒性有关。但是多柔比星导致心肌细胞凋亡的具体机制目前尚未研究透彻。
众多研究表明,多柔比星作用心肌细胞后,细胞内产生的大量氧自由基在细胞凋亡过程中发挥重要作用[9-10]。这在我们前期的实验也得到证实[11]。本实验通过选取多柔比星0.1、0.3、1、3 和10 μmol/L 5个浓度梯度以观察其对心肌细胞凋亡的影响。我们发现,多柔比星作用心肌细胞后,硝基化修饰的Trx明显增加。Trx 硝基化后可导致其活性降低甚至丧失,抑制ASK1 的能力下降,进而导致ASK1 介导的凋亡的发生[12-14]。因此,Trx 的硝基化导致活性丧失可能是多柔比星导致心肌细胞凋亡的机制之一。过氧亚硝基阴离子被认为是蛋白硝基化修饰的重要因素[4],我们选取了过氧亚硝基阴离子清除剂MnTMPyP作为保护剂,对Trx 的硝基化以及Trx-ASK1 等凋亡相关信号分子进行进一步探讨。实验证实MnTMPyP 可以降低多柔比星诱导的心肌细胞凋亡,细胞内的Trx硝基化形式含量显著降低,caspase-3 和PARP-1 蛋白发生活化形式降低。降低Trx 硝基化可能是多柔比星诱导心肌细胞凋亡的途径之一,多柔比星导致的Trx硝基化可能与细胞内产生ONOO-有关。
Trx 是细胞内一种重要的二硫键还原酶,对维持细胞内蛋白质的氧化还原状态起着非常重要的作用。生物体内的Trx 系统包括Trx、Trx 还原酶(thioredoxin reductase,TrxR)、还原型辅酶II(NADPH)和Trx 过氧化物酶(thioredoxin peroxidase,TrxP),这是一个控制细胞内氧化还原平衡状态的重要的氧化还原酶系统。Trx 除了具有抗氧化作用外,还具有直接抗凋亡作用,其作用机制与细胞内的ASK1 密切相关[12-13]。ASK1 是丝裂原激活蛋白激酶激酶激酶家族(MAPKKK)中的一员,在凋亡信号中发挥着重要作用。p-ASK1 (Ser83)、p-ASK1(Ser967)发生去磷酸化或者p-ASK1(Thr845)发生磷酸化均可代表ASK1 的活化。ASK1 活化可以激活丝裂原活化蛋白激酶激酶(MAPKK)家族中的MAPKK4、MAPKK7、MAPKK3 和MAPKK6,进而活化丝裂原活化蛋白激酶(MAPK)家族中c-Jun 氨基端激酶(JNK)和p38 这2 个促凋亡蛋白激酶。促凋亡蛋白激酶p38 与JNK 的活化会通过caspase 依赖途径激活caspase-3 和PARP-1 的剪切致使PARP-1 失活导致细胞失去重要的DNA 修复功能而发生凋亡。还原型Trx 主要是通过Cys-32、Cys-35 的巯基与ASK1的N 端连接,通过抑制了ASK1 活性进而抑制细胞凋亡。我们以前的实验已经证实给予多柔比星后,Trx 与ASK1 发生了分离可能是导致心肌细胞凋亡原因之一。多柔比星心脏毒性研究中发现,心肌细胞中蛋白质的硝基化形式明显增加[4,15]。Trx 含有的酪氨酸残基,在ONOO-大量产生时也可以发生硝基化修饰。Trx 硝基化失活是否参与多柔比星导致的凋亡,目前没有详细报道。对此,我们用多柔比星刺激心肌细胞进行实验,免疫沉淀和免疫印迹法检测Trx 硝基化,发现多柔比星刺激心肌细胞后,细胞内Trx-NT 表达明显增加;Trx-ASK1 复合形式含量减少(P <0.05);利用p-ASK1(Ser967)抗体进行Western blotting 检测,p-ASK1(Ser967)表达显著减少(P<0.05),ASK1 发生了明显活化;p-p38 MAPK 表达显著增加(P <0.05)。多柔比星体外诱导心肌细胞凋亡过程中Trx-ASK1 分离增多,ASK1 活化,导致细胞凋亡的发生,这可能与Trx 硝基化修饰有关。使用ONOO-清除剂MnTMPyP 降低心肌细胞凋亡的过程中发现,Trx 硝基化程度减轻,Trx-ASK1 调节的凋亡相关信号分子受抑制,说明ONOO-清除剂MnTMPyP可以抑制多柔比星导致的Trx 硝基化修饰的增加。本文主要是探讨多柔比星诱导心肌细胞凋亡过程中Trx 的硝基化水平与凋亡的关系。对于这一过程Trx硝基化的具体机制和途径有待进一步探讨。综上所述,本实验初步证实多柔比星诱导心肌细胞凋亡过程中,Trx 硝基化修饰可能是多柔比星诱导心肌细胞凋亡的机制之一,这可能与多柔比星刺激ONOO-的产生有关。本文探讨的Trx 硝基化失活对凋亡的影响,将有助于进一步探讨多柔比星心脏毒性以及心肌细胞凋亡具体机制的研究。
[1] Carvalho C,Santos RX,Cardoso S,et al. Doxorubicin:the good,the bad and the ugly effect[J]. Curr Med Chem,2009,16(25):3267-3285.
[2] Zhang YW,Shi J,Li YJ,et al. Cardiomyocyte death in doxorubicin-induced cardiotoxicity[J]. Arch Immunol Ther Exp (Warsz),2009,57(6):435-445.
[3] Nuriel T,Hansler A,Gross SS. Protein nitrotryptophan:formation,significance and identification[J]. J Proteomics,2011,74(11):2300-2312.
[4] Mukhopadhyay P,Rajesh M,Batkai S,et al. Role of superoxide,nitric oxide,and peroxynitrite in doxorubicin-induced cell death in vivo and in vitro[J]. Am J Physiol Heart Circ Physiol,2009,296(5):H1466-H1483.
[5] Yin T,Hou R,Liu S,et al. Nitrative inactivation of thioredoxin-1 increases vulnerability of diabetic hearts to ischemia/reperfusion injury[J]. J Mol Cell Cardiol,2010,49(3):354-361.
[6] Tao L,Gao E,Hu A,et al. Thioredoxin reduces post-ischemic myocardial apoptosis by reducing oxidative/nitrative stress[J]. Br J Pharmacol,2006,149(3):311-318.
[7] Tao L,Jiao X,Gao E,et al. Nitrative inactivation of thioredoxin-1 and its role in postischemic myocardial apoptosis[J]. Circulation,2006,114(13):1395-1402.
[8] Simpson P,Savion S. Differentiation of rat myocytes in single cell cultures with and without proliferating nonmyocardial cells. Cross-striations,ultrastructure,and chronotropic response to isoproterenol[J]. Circ Res,1982,50(1):101-116.
[9] 吴伟康,杨 辉,赵明奇. 阿霉素性心力衰竭模型的氧化应激和凋亡机制[J]. 中国病理生理杂志,2004,20(8):1437-1439.
[10]Gao J,Xiong Y,Ho YS,et al. Glutathione peroxidase 1-deficient mice are more susceptible to doxorubicin-induced cardiotoxicity[J]. Biochim Biophys Acta,2008,1783(10):2020-2029.
[11]黄海高,李悦山. Trx-ASK1 在多柔比星诱导的乳鼠心肌细胞凋亡中的作用[J]. 中国病理生理杂志,2012,28(6):1028-1033.
[12]Nishida K,Otsu K. The role of apoptosis signal-regulating kinase 1 in cardiomyocyte apoptosis[J]. Antioxid Redox Signal,2006,8(9-10):1729-1736.
[13]Lu J,Holmgren A. Thioredoxin system in cell death progression[J]. Antioxid Redox Signal,2012,17(12):1738-1747.
[14]陶 凌,臧益民,刘慧荣,等. 硝基化修饰导致硫氧还蛋白不可逆的丧失了对心脏的保护作用[J]. 心脏杂志,2005,17(4):307-312.
[15]Pacher P,Liaudet L,Bai P,et al. Potent metalloporphyrin peroxynitrite decomposition catalyst protects against the development of doxorubicin-induced cardiac dysfunction[J]. Circulation,2003,107(6):896-904.
