四尾栅藻对铜绿微囊藻光合活性及毒素产生与释放的影响
2013-12-21刘文清赵南京段静波王志刚张玉钧殷高方石朝毅
方 丽, 刘文清, 赵南京, 段静波, 王志刚,2, 肖 雪, 张玉钧, 刘 晶, 殷高方,石朝毅
(1. 中国科学院环境光学与技术重点实验室 中国科学院安徽光学精密机械研究所, 合肥 230031;2. 扬州大学扬子津校区环境学院,扬州 225127)
湖泊富营养化日益加剧,随之而来的蓝藻水华成为全球关注的热点环境问题。水华暴发会进一步加剧水体污染,带来更严重的危害,如造成水体恶臭,产生毒素等,给饮用水处理造成困难。铜绿微囊藻(Microcystisaeruginosa)是全球蓝藻水华的常见藻种之一,属于蓝藻门蓝藻纲微囊藻科。细胞结构简单,为单细胞个体或群体,群体种类在细胞壁外分泌果胶类物质构成胶质鞘膜[1]。它产生的微囊藻毒素(MCs)是一类具有生物活性的环状七肽化合物,对哺乳动物、鱼类、鸟类以及无脊椎动物皆有毒性[2]。据报道,微囊藻毒素有60多种同分异构体,其中微囊藻毒素LR(MC-LR)就是最常见的三种之一。长期以来,各国学者对铜绿微囊藻进行了大量的研究报道,包括其生长、产毒和环境影响因子等方面。四尾栅藻(Scenedesmusquadricanda)隶属于绿藻门绿藻纲绿球藻目栅藻科,为不运动的单细胞,通常由2、4或8个细胞组成。已有报道从竞争的角度研究铜绿微囊藻的产毒机理。许秋瑾[3]等的研究结果表明存在竞争时,铜绿微囊藻比单种培养时产生更多的毒素。万蕾等[4]研究了不同营养条件下,铜绿微囊藻与四尾栅藻的竞争实验。结果表明在贫营养水平下栅藻能够刺激微囊藻生长,富营养水平下,竞争抑制作用与N/P 有关,在较低氮磷浓度的水体中,微囊藻容易成为优势种,而在较高氮磷浓度的水体中,四尾栅藻更容易成为优势种。张坤等[5]研究了5种微藻对铜绿微囊藻生长的影响。结果表明5种微藻的初始密度对铜绿微囊藻的生长有显著影响。当铜绿微囊藻初始密度较小时,微藻密度增加对铜绿微囊藻生长抑制效果越明显,而当铜绿微囊藻的初始密度较高时,5种微藻密度的增加对铜绿微囊藻生长抑制效果不明显。Fogg等[5]研究发现藻类在生长发育过程中会不断向周围环境中释放碳水化合物、酶、脂类、抑制或促进因子、毒素等来影响水体中其他藻类的生长发育。Piazzi 等[6]研究了2种微藻种内和种间竞争,发现在细胞密度较低时,2种微藻竞争不明显,随着培养时间的增加,细胞密度变大,微藻间的抑制作用变得明显,培养后期的竞争可能是因为空间和营养资源缺乏。
由于藻类是光合自养生物,光合作用很重要,藻类各种结构和生理过程都影响生长,但与生长关系最密切的是光合作用[7],研究藻类生物量与光合作用活性之间的关系,通过监测藻类荧光预知其生长潜能,对水华预警有重要意义。本实验选择产MC-LR的铜绿微囊藻与不产该种毒素的四尾栅藻为实验藻种,进行了为期58天的检测分析,研究了四尾栅藻对铜绿微囊藻毒素MC-LR产生与释放的影响,以及光合作用活性与生物量的相关性,为进一步研究野外混合藻的生长与产毒打下基础,以及为通过活性预警蓝藻水华提供基础理论依据。
1 材料与方法
1.1 仪器与试剂
实验分析过程中用到的仪器主要包括:防交叉干扰自动固相萃取装置(美国sigma公司); C18固相萃取小柱(500 mg/3 mL supelco En-vi小柱);Agilent 1200高效液相色谱仪;Eclipse XDB-C18色谱柱;水样荧光仪(WATER-PAM,walz,Germany);HP400G型智能光照培养箱(武汉瑞华仪器设备有限责任公司);Molecular超纯水机;不锈钢手提式压力蒸汽灭菌器;迷你水浴锅(W1106-230);H-1650R型台式高速冷冻离心机;NMT-2800氮吹仪(天津奥特塞恩斯);细胞计数仪CounterStar(上海睿钰生物科技有限公司)。
实验所用试剂主要有:甲醇(色谱纯);三氟乙酸(色谱纯);去离子水;纯净水;磷酸盐缓冲溶液(0.05 mol/L);
标准样品的配制:MC-LR标准样品购于北京伊普瑞斯公司。用20%甲醇(分析纯)将标准毒素配制成50、20、10、5、2、1、0.5、0.2、0.1、0 μg/mL,采用HPLC进行分析,以测量标准样品谱线峰面积与其对应的浓度制作标准曲线,结果如图1所示。
液相色谱条件:色谱柱温度40 ℃;流动相为甲醇与磷酸盐缓冲溶液按体积比57∶43混合;检测器为二极管阵列检测器波长238 nm;流速为1 mL/min,进样量为20 μL[8]。
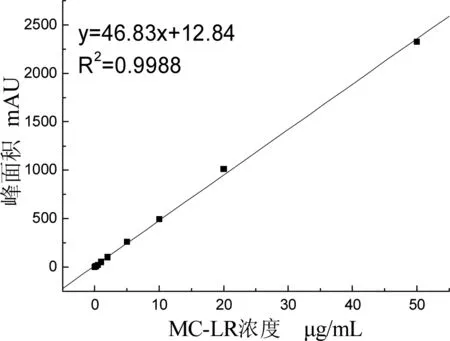
图1 微囊藻毒素LR标准曲线
1.2 实验材料
实验所用铜绿微囊藻(FACHB-905)及四尾栅藻(FACHB-1297)购自中科院水生所。取60个灭菌的250 mL锥形瓶,分别加入130 mL已灭菌的BG-11培养基及3 mL(2.5×105个/mL)对数生长期的铜绿微囊藻。分为A,B两组,每组30瓶 ,向B组锥形瓶中分别加入3 mL(2.5×105个/mL)对数生长期的四尾栅藻。两组藻在培养箱中随机放置,每天摇动一次并随机交换位置。在光照度3000 lx、温度为25 ℃、光暗比为14 h/10 h下培养(在预实验中,该条件下四尾栅藻及铜绿微囊藻在BG-11培养基中均能很好的生长)。每隔2 d测定藻细胞数、胞外毒素MC-LR、胞内毒素MC-LR及光合活性,每次2个平行样,后期藻生长缓慢,采样间隔适当增大。
1.3 藻细胞计数
分别取1 mL样品于250 mL锥形瓶中,用去离子水稀释至100 mL,采用细胞计数仪分别计数两种藻活细胞个数。
1.4 胞外毒素MC-LR测定
分别取100 mL藻液,离心分离藻细胞,上清液过0.45 μm玻璃纤维醋酸酯膜后固相萃取,依次用10 mL纯水、10 mL20%甲醇淋洗,再用5 mL 90%甲醇(含0.1%三氟乙酸)洗脱[9]。洗脱液在40 ℃水浴中氮气吹干,1 mL甲醇将吹干物转移至HPLC进样瓶中(分两次,每次0.5 mL)。再次氮气吹干,用50%甲醇定容至200 μL,待测。测定结果以藻液体积归一化,转化成锥形瓶中的毒素浓度。计算公式如下:

其中,c为锥形瓶中毒素MC-LR浓度,单位μg/L;X为测得的色谱进样瓶中的MC-LR浓度,单位μg/mL;0.2为定容体积200 μL;100为藻液体积100 mL。
1.5 胞内毒素MC-LR测定
上述离心后的藻细胞用去离子水转移至锥形瓶中,沸水浴处理20 min[10-11],提取毒素,离心,取上清液,残渣用去离子水洗涤一次,离心,合并上清液。其余处理同细胞外毒素。为了方便对比,最终以藻液体积归一化内毒素水平,单位为μg/L。
1.6 光合活性测定
藻的光合作用活性以叶绿素a荧光表示。叶绿素荧光分析技术利用植物体内叶绿素作为天然探针,研究和探测细胞光合生理状况及各种外界环境因子对其细微影响[12]。Yield值为光系统Ⅱ(PSⅡ)实际光化学量子效率,即实际光合效率,它反映PSⅡ反应中心在有部分关闭情况下的实际原初光能捕获效率,细胞不经过暗适应在光下直接测得:
Yield=(Fm′-F)/ F′
其中,F′—光下最大荧光,光适应下全部PSⅡ中心都关闭时的荧光强度
F—实际荧光产量,仪器测定时F值为打开饱和脉冲前的荧光
经暗适应后测得的Yield为PSⅡ最大光合效率:
Yield=(Fm-F0)/Fm
其中,Fm—最大荧光,已经暗适应的光合机构全部PSⅡ中心都关闭时的荧光强度
F0—最小荧光或初始荧光,已经暗适应的光合机构全部PSⅡ中心都开放时的荧光强度
各参数由水样荧光仪(WATER-PAM,Walz,Germany)测定[9]。为减小误差,测定时间均在9点钟左右,于室温下暗适应5 min后测定F0及Fm,计算得出Yield值。
2 结果与讨论
2.1 四尾栅藻对铜绿微囊藻生物量及活性的影响
A、B组铜绿微囊藻生物量如图2所示。培养前12 d,A、B组藻细胞生物量均呈对数增长,之后缓慢增长,数量级不变。达到最大值后,由于营养盐的限制,藻生长受到抑制,生物量开始下降。几乎在整个生长周期中,A组铜绿微囊藻生物量均大于B组。这与许秋瑾等的研究结果一致[3],是因为B组四尾栅藻与铜绿微囊藻竞争营养盐及生长空间,抑制了藻的大量增殖。A组生物量约在第38 d达到最大值,B组提前8 d左右达到最大值。
A、B组藻的光合作用活性如图3所示。利用Water-PAM测定了两组藻yield值,其中B组为铜绿微囊藻与四尾栅藻混合藻液的yield值。yield值是PSⅡ最大光化学量子产量,反映了植物对光量子的最大利用潜能,非胁迫条件下该参数的变化极小,不受物种和生长条件的影响,胁迫条件下该参数明显下降,是用来研究植物受到胁迫对光合作用影响的重要指标[13,14]。由图可见,培养前期两组藻yield值不断增大,第12 d达到最大。结合生物量数据来看,前12 d生物量呈对数增加,即对数期藻光合作用活性较强,藻生长潜能大。随后yield值开始逐渐下降,生物量呈稳定增加,数量级达到最大,不再增加。从开始培养至测量结束,B组yield值均高于A组,前期二者差距较小,随着培养时间的增加,差距也在增大。58 d内,平均高出0.037。分别培养蓝藻和绿藻时,蓝藻的yield值一般较低,符合蓝藻的生理特点,绿藻则较高[14]。本实验中B组yield值始终高于A组,就是因为B组中存在绿藻。目前无法分开测定混合藻中铜绿微囊藻的光合作用活性。关于光合作用活性的研究多集中在环境影响因子、营养盐限制等方面,种间竞争对光合作用活性的研究几乎还是空白。本实验结果初步证明,藻类生长过程中,光合作用活性与生物量之间存在一定的相关性,活性大,藻生长迅速,活性小,则生物量增加缓慢,数量级不变。监测自然水体中藻类光合作用活性,对蓝藻水华有预警作用,值得更加系统深入的研究。
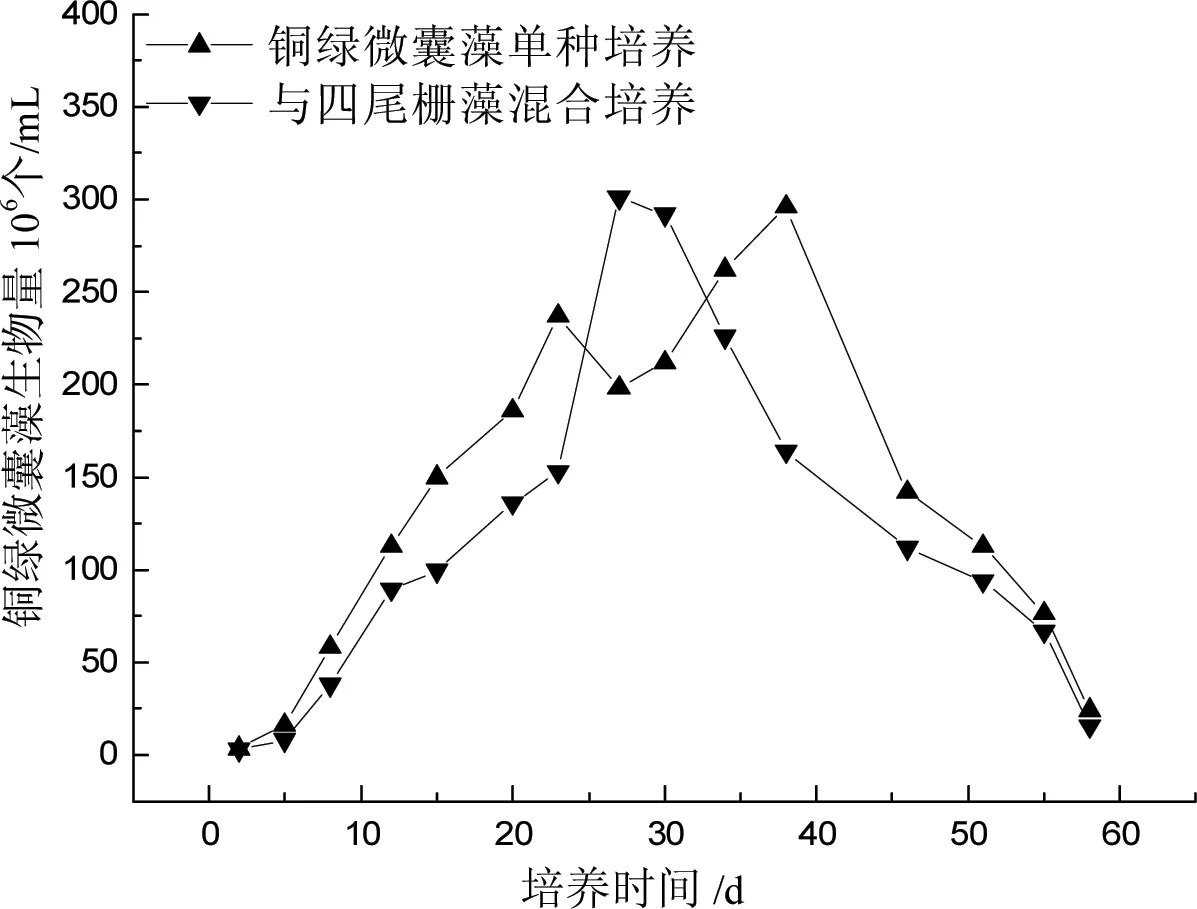
图2 四尾栅藻对铜绿微囊藻生物量的影响

图3 四尾栅藻对铜绿微囊藻光合活性的影响
2.2 四尾栅藻对铜绿微囊藻胞外毒素MC-LR的影响
A、B两组藻液的细胞外MC-LR毒素浓度如图4所示。A组藻液胞外MC-LR毒素始终处在较低水平,总体呈缓慢上升趋势。因为此次实验,营养盐较丰富,整个实验周期中,藻生长尚未达到衰亡期,细胞没有开始大量死亡,释放毒素较缓慢。而B组培养38 d后,胞外毒素浓度上升加快,说明此时藻已开始大量死亡。由图可见,B组毒素浓度始终高于A组,培养后期B组上升速度加快,导致两者差距越来越大,约第55 d达到最大值,此时B组胞外毒素比A组高606.7 μg/L。由此得出,当起始浓度相同时,铜绿微囊藻单种培养比与四尾栅藻共培养释放出更少的毒素MC-LR,这是由于四尾栅藻的存在,增加了铜绿微囊藻的生存压力,释放出更多的毒素,从而在竞争中占据优势。同时四尾栅藻也会分泌某些物质抑制铜绿微囊藻的生长。有研究表明某些真核藻分泌的化感物质可有效抑制铜绿微囊藻的生长和光合作用,具体影响有待更深入的研究。一般认为微囊藻毒素为细胞内毒素,在细胞内合成,细胞死亡破裂后释放到环境中。但是已有研究发现藻类在死亡之前也会向水体中释放毒素,相关研究报道还不多,尚无明确结论。
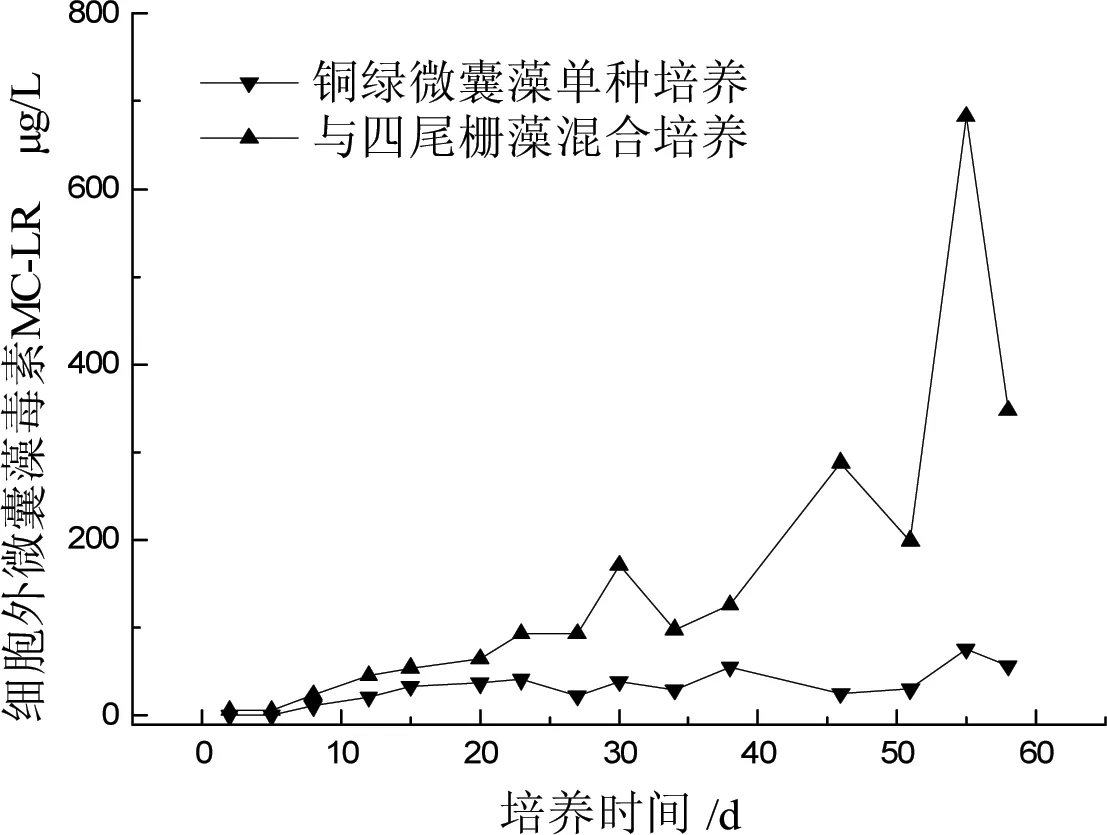
图4 四尾栅藻对铜绿微囊藻胞外毒素MC-LR的影响
2.3 四尾栅藻对铜绿微囊藻胞内毒素MC-LR的影响
A、B 两组藻整个生长过程中细胞内毒素MC-LR浓度水平见图5。其中A组总体呈上升趋势,38 d达到最大值78.53 μg/L,随后开始下降。B组胞内MC-LR浓度也始终高于A组,最大值出现在第46 d,为334.35 μg/L。后期与A组距离逐渐拉大。
关于微囊藻毒素产生机理的研究是目前的热点,有很多假设,但还无令人满意的结果。有两种可以考虑的可能性,一是遗传差异即微囊藻有毒株和无毒株的遗传结构不同,另一种是环境因子影响或改变毒性[15]。本文研究表明,存在竞争时,铜绿微囊藻产毒会发生变化。以往已有研究表明,混合培养比单种培养产生更多的毒素[3]。本研究结果与此一致,A组生物量较B组大,但细胞内外毒素MC-LR均低于B组。随着培养天数的增加,细胞内毒素水平上升。达到最大值后逐渐下降。日本学者Watanabe等人的研究表明,铜绿微囊藻停止生长后,在一段时间内存在于固形物(包括藻和其他微生物)中的藻毒素含量与存在于水体中的藻毒素有很好的消长关系[16]。直至细胞全部死亡后,固形物中的毒素为零而水体中毒素达到最大,随后开始被色素、光、微生物等降解,浓度开始下降。本次实验中A、B两组藻均遵循这个规律。参考细胞生物量数据,本研究还发现,A组铜绿微囊藻个数总是比B组多,由此再结合细胞内、外毒素MC-LR浓度可初步推断,在四尾栅藻存在的情况下,每个铜绿微囊藻细胞会产生更多的毒素,且释放出比单种培养更多的毒素到水体中。关于单个藻细胞产毒量的研究报道还不多,有待进一步更精确的研究。

图5 四尾栅藻对铜绿微囊藻胞内毒素MC-LR的影响
3 结论
本实验初步得出以下结论,铜绿微囊藻与四尾栅藻共培养时,生长受到一定的限制,最大生物量提前,单个铜绿微囊藻细胞比纯种培养时产生更多的毒素。Yield值上升时,生物量呈对数增加,藻生长潜能大,暴发蓝藻水华可能性大。Yield值达到最大,则生物量数量级也达到最大,不再增加。通过在线监测藻类荧光,可使预警蓝藻水华成为可能。实验初步研究了实验现象,关于机理还需更加深入的研究。共培养时,不仅生物量、毒素产生与释放、光合作用活性等发生了变化,藻细胞尺寸大小的变化也同样反应了藻体特征参数的改变,而关于这方面的研究报道还很少,有待完善。
[1]栗越妍.铜绿微囊藻和四尾栅藻的生长及竞争特性研究[D].中国环境科学研究院,2009.
[2]Penaloza R,Rojas M,Vila I,et al. Toxicity of a soluble peptide from microcystis sp to zooplankton and fish[J]. Freshwater Biol,1990,24:233-240.
[3]许秋瑾,高 光,陈伟民.从种群竞争的角度初步研究微囊藻的产毒机理[J].生物学杂志,2004,21(1):16-18.
[4]万 蕾,朱 伟,赵联芳.氮磷对微囊藻和栅藻生长及竞争的影响[J].环境科学,2007,28(6):1230-1235.
[5]张 坤,戴习林.5种微藻及其密度对铜绿微囊藻生长的影响[J].广东农业科学,2012,10:166-178.
[6]Muller-Feuga A. The role of microalgae in aquaculture: situation and trends[J]. J Appl Phycol,2000,12(3/5):527-534.
[7]贾晓会,施定基,史锦红,等.巢湖蓝藻水华形成原因探索及“优势种光合假说”[J].生态学报,2011,31(11):2968-2977.
[8]中华人民共和国国家标准.GB-T 20466-2006水中微囊藻毒素的测定.
[9]胡智泉,刘永定,肖 波.微囊藻毒素对几种淡水微藻的生长和光合活性的影响[J].生态环境,2008,17(3):885-890.
[10]李现尧,刘晓文,史全良.一株微囊藻毒素降解辅助菌的分离和鉴定[J].微生物学通报,2010,37(3):473-478.
[11]Metcalf J S,Codd G A. Microwave oven and boiling waterbath extraction of hepatotoxins from cyanobacterial cells[J]. FEMS Microbiology Letters,2000,184:241-246.
[12]李 娜.CO2浓度与铜绿微囊藻生长关系的研究[D].哈尔滨工业大学,2010.
[13]张守仁.叶绿素荧光动力学参数的意义及讨论[J].植物学通报,1999,16(4):444-448.
[14]吴晓东,孔繁翔,曹焕生,等.越冬浮游植物光合作用活性的原位研究[J].湖泊科学,2007,19(2):139-145.
[15]李效宇,宋立荣,刘永定.微囊藻毒素的产生、检测和毒理学研究[J].水生生物学报,1999,23(5):517-523.
[16]徐立红,周炳升,张甬元,等.微囊藻毒素环境行为的初步研究[J].水生生物学报,1997,21(1):85-89.
