长白山北坡不同林型内红松年表特征及其与气候因子的关系
2013-12-19高露双张思行赵秀海
陈 列,高露双,张 赟,张思行,赵秀海
(北京林业大学省部共建森林培育与保护教育部重点实验室,北京 100083)
自20世纪以来中国东北地区升温明显,1900—1999间温度升高了2℃[1-2]。同时,东北东部地区的降水变化也较为剧烈[3]。温度和降水等气象因子的变化对树木的生长有着重要影响。树木年轮记录了树木自身生长与外界环境变化的响应[4]。以往研究表明树轮宽度与气象因子有着明显的相关关系[4-6]。利用树木年轮可以从时间和空间尺度上探讨气候变化对树木生长的影响,从机理上解释环境变化影响树木生长的原理。在全球气候发生明显变化的背景下,利用树轮资料研究树木生长与气象因子的关系显得更为重要。
长白山地区气候变化剧烈,是我国东部开展树轮研究较早的区域之一[7]。其保存完好的高山垂直植被带是我国东北地区森林生态系统的典型代表,是研究气候与树木生长关系的理想地区。以往该地区的树轮研究多集中于高海拔(1300m以上)的针叶林[8-9]或林线位置(2000m)的岳桦林[10],而低海拔地区的树轮研究相对较少[11]。阔叶红松林是长白山地区水平地带性植被,分布于海拔500—1100m,红松是该森林类型的建群种。前人已对上下限红松的生长与气候的关系做了探讨,取得了一定的成果[12-14]。但对于适合红松生长的海拔区域内红松生长的气候响应机制研究还很少。
本文以树木年代学的方法探究了长白山北坡中海拔地区不同林型内建群种红松径向生长对气候变化的响应,是对该地区已有研究的补充,为预测未来气候变化如何影响红松生长提供依据。
1 材料和方法
1.1 研究区概况
长白山位于我国东北地区吉林省东南部的中朝交界处,北纬 41°23'—42°36',东经 126°55'—129°08',是我国乃至全球自然生态系统保存最完整的地区之一。本地区气候属于受季风影响的温带大陆性气候,具有冬季寒冷而漫长,夏季温暖多雨而短暂的特点。全区水热状况的特点是水量充沛,热量不足,湿润状况良好。年平均气温为1.18℃,年累积降水量为600—1340mm。降水多集中在夏季,6—9月份降水量占全年降水量的80%。年相对湿度在70%左右。
长白山地区红松的垂直分布一般在海拔500—1200m之间,单株红松可达1400m。红松喜好温和湿润的气候条件,对温度的适应幅度较大。红松喜光,耐寒力强,不耐干旱,喜湿润、土层深厚、肥沃、排水和通气良好的微酸性土壤。
1.2 样本采集
2008年,在杨桦红松林(H1)和椴树红松林(H2)内,分别设置5个20m×20m样方,选择样方内林冠层的红松,在胸高处(距地面130cm)用生长锥钻取树芯,每株树锥取两个树芯样本。取样方向与山坡坡向垂直,以避免坡度的影响造成年轮挤压现象。采样点内红松应至少达到30棵,若样方内数量过少,无法满足样本量要求,可在样方附近随机补取样木。采样点概况见表1。

表1 采样点基本情况Table 1 Description of sampling sites
1.3 年表建立
样芯采回后固定,风干和打磨以达到树木年轮学分析的要求,然后测量年轮宽度并交叉定年。本文运用LinTab5年轮分析仪器在0.001mm水平上测量树轮宽度。为保证测量和定年的准确性,用COFECHA程序对交叉定年作进一步检验[15],剔除问题较大的树芯。最终保留H1样点的26棵树50个样芯,H2样点的27棵树52个样芯。
本论文运用ARSTAN软件,采用步长35a的样条函数法消除与树龄相关的生长趋势(简称去趋势),并对去趋势的序列以双重平均法合成标准年表(STD)。采样点森林郁闭度高,树木间竞争激烈,为消除竞争对树木生长的影响,又以时间序列的自回归模型对去趋势的序列进行再次标准化,以双权重平均法将差值序列合成差值年表(RES)。由于RES的统计特征要好于STD(表2),因此本文用RES代替红松的径向生长来分析与气候的响应关系。

表2 红松标准化和差值年表统计特征及公共区间(1930—2008)分析Table 2 Statistic characteristics and common interval(1930—2008)analysis of standard and residual chronologies of Pinus koraiensis
1.4 气候资料来源
本研究选取了长白山地区松江气象站点的月平均温度和月降水量2种气候要素。采用Kendall[16]和Double-mass方法[17]检验对气象站气候资料进行均一性检验,结果表明气候资料无随机突变和明显不均匀分布情况,可用来代表自然气候的变化。
1.5 数据分析
本文采用响应函数来分析树木生长对气候的响应关系,统计过程应用树轮研究学专用软件DENDROCLIM2002程序来实现,显著水平P<0.05[18]。以往研究表明树木生长受到当年和上一年气候因子的影响[4],故本文选取上一年6月到当年9月的逐月气候变量(包括月平均温度和月降水量)与树轮标准化年表进行响应函数分析。同时还采用特征年分析[15]的方法来探究造成宽轮和窄轮的气候因素。
2 结果
2.1 年表的基本统计特征
平均敏感度(MS)是年轮宽度逐年变化状况,主要反映气候的短周期变化或高频变化。H1和H2的平均敏感度均较高(表2),说明红松生长对气候变化敏感。从图1可以看出,椴树红松林(H2)年轮宽度的波动幅度更大,说明H2内的红松更适合于做年轮气候学的分析。两个样点树与树之间的相关系数均达到0.6(表2),说明树木径向生长很好的反映了气候变化的一致性。另外,信噪比,样本的总体代表性和第一主成分所解释的方差量都比较高(表2),进而证实了红松适合于年轮气候学研究。
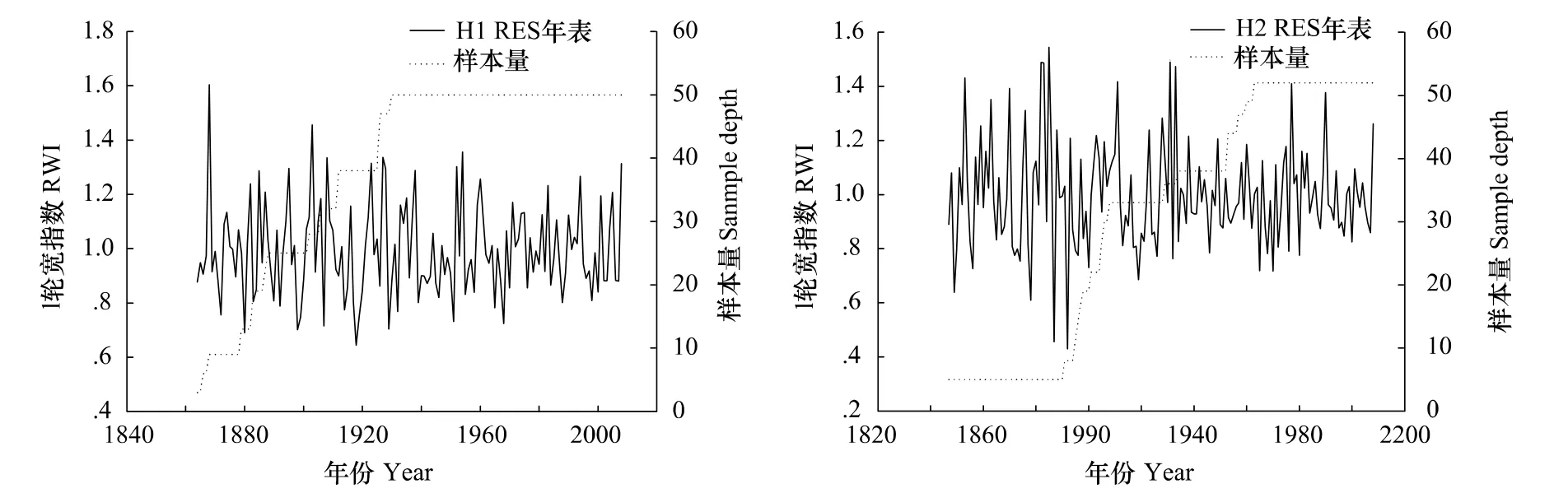
图1 H1和H2样地内红松差值年表Fig.1 Residual chronologies of Pinus koraiensis in poplar-birch(H1)and linden-Korean pine forest(H2)
2.2 径向生长对气候因子的响应
在杨桦红松林内(H1),红松的年轮宽度和平均温度没有显著的相关关系(图2),红松的年轮宽度和当年7月与上一年9月的降水呈显著正相关关系(图2)。说明在杨桦红松林,温度不是影响红松生长的主要因子,红松的径向生长主要受当年生长季以及上年生长季末降水的影响。在椴树的阔叶红松林内(H2),红松的年轮宽度与当年3、4月份的平均气温呈显著正相关(图3),说明生长季前的升温对红松生长有积极影响。降水方面,红松的年轮宽度和当年的6、7月呈显著正相关(图3),说明生长季的降水对红松生长有着重要影响。同时,红松的年轮宽度也与上一年9月的降水呈显著正相关,与上一年7月的降水呈显著负相关(图3)。
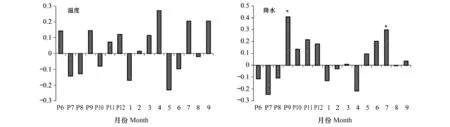
图2 杨桦红松林内红松差值年表与气候因子的关系Fig.2 Response function analysis for the relationship between residual chronology of P.koraiensis and climate factors in poplar-birch Korean pine forest

图3 椴树红松林内红松差值年表与气候因子的关系Fig.3 Response function analysis for the relationship between residual chronology of P.koraiensis and climate factors in linden-Korean pine forest
2.3 特征年分析
在有气候数据记录的1960—2008年间,杨桦红松林(H1)样点内红松在1968年出现了极窄年份,当年生长量为0.735mm,较历年平均年生长量降低了0.4mm。通过分析极端年份的气候因子特征,发现1968年当年7月和上一年9月的降水量均低于历史平均水平,差值分别为373mm和570mm(图4)。同时,1968年7月份的温度也低于历史平均水平(图4)。这说明当年7月的低温和降水异常,以及上一年9月的降水偏少是产生窄轮的主要原因。椴树红松林(H2)的红松则在1970年表现出极窄的轮宽,仅为0.619mm,较历年平均年生长量降低了0.38mm。1970年3月份的月平均气温低于历史平均水平(低7℃)(图5)。在降水方面,无论是在红松生长旺盛期的当年7、8月份,还是在红松生长末期的上一年9月,1970年的月平均降水量均低于历史平均水平(图5)。

图4 降水量单年分析(1968年、1970年)距平分析Fig.4 Single year analysis of monthly precipitation in 1968 and 1970
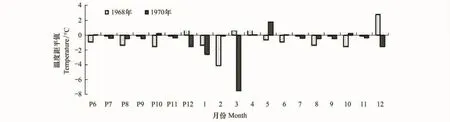
图5 温度单年分析(1968年、1970年)距平分析Fig.5 Single year analysis of monthly temperature in 1968 and 1970
3 讨论
3.1 径向生长与气候因子的关系
杨桦红松林内红松的径向生长主要受降水影响。7月是红松的生长旺季,也是降水最多的月份,高温高湿的气候条件十分有利于树木的光合作用,生产出足够的光合产物促进树木的生长[19]。反之,7月的温度最高,蒸发量相对较大,降水不足便会抑制树木的径向生长[4]。9月处于生长季末期,树木仍能进行一定的光合作用,丰富的降水有利于红松光合产物的积累以及土壤水分的涵养,为来年树木的生长提供充足的养分和良好的条件[20-21]。生长季末期降水的“滞后效应”在其它针叶树种上也有所体现[22]。
椴树红松林内红松的径向生长受温度和降水共同影响。初春(3和4月)温度升高促进了形成层活动提早,有利于树木及早打破休眠,从而延长了生长季,有利于树木的生长[23]。另外,春季气温升高加速林地积雪的融化和地温的升高,有利于根系活动,从而促进了树木的生长[24]。其他研究表明,同为五针松的华山松也受到初春温度的重要影响[25-26]。同杨桦红松林类似,椴树红松林内红松的年轮宽度与当年生长季(6和7月)以及上一年生长季末(9月)的降水呈显著正相关。同时,上年7月的降水成为当年红松生长的限制因子。这可能是因为7月是红松生长最旺盛的时期,充足的降水大大促进了树木的光合作用,消耗了过多的光合产物,减少了营养物质的积累,从而影响到来年树木的生长。陈力等[12]研究也发现,红松生长与上年7月的降水显著负相关。上一年生长季优越的气候条件限制来年树木的生长,这一现象在其他树木年轮研究中也有所体现[27]。
本研究表明,无论是在杨桦或椴树红松林内,当年生长季和上一年生长季末的降水对红松的径向生长均有着重要影响,这与前人的研究结果一致。例如,高露双等[13]发现红松的径向生长与当年7月的降水呈显著正相关;Yu等[28]的研究结果也表明,生长季的丰富降水促进了红松的径向生长;李广起等[14]发现上一年生长后期(8和9月)的降水与红松年轮宽度有着明显的正相关关系。长白山地区虽然降雨量较大,但是降水仍然是影响红松生长的主要因子,这可能与红松自身的喜湿特性密切相关。
杨桦红松林内红松生长仅与降水显著相关,而在椴树红松林内,温度也成为影响红松生长的主要因子。这种差异可能与两样点所处的海拔相关。以往研究表明,降水是分布下限树木生长的限制因子[29-30],而分布上限树木的生长则受低温限制[31-32]。杨桦红松林所处海拔较低,温度条件适宜,故红松对气候因子的响应只反映在降水上。椴树红松林所处海拔较高,接近于红松分布的上限,该样点的温度环境更寒冷,因此红松对温度更为敏感。温度的升高,尤其是春季温度的升高,延长了生长季,有利于红松的早期生长。Yu等[28]研究发现,生长在上限的红松与3月和4月的温度呈显著正相关。同时,其他研究也表明上限红松的径向生长与生长季的温度呈明显的正相关关系[13-14]。
3.2 特征年对径向生长与气候因子关系的验证
通过选取有代表性的年份分析气候因子对树木生长的影响,从而验证前面响应函数的相关分析结果。特征年分析结果表明无论在杨桦(H1)或椴树(H2)红松林内,当年7月和上一年9月的降水偏少是引起窄轮的主要原因。从而进一步验证了响应函数所得到的结果,即当年生长季以及上一年生长季末的降水是决定红松年轮宽度的主要因素。结果同时指出,生长季的温度(杨桦红松林)和初春温度(椴树红松林)偏低也是形成窄轮的重要原因。这也和本文响应函数以及前人的研究结果相一致。生长季充足的降水和适宜的温度使得红松生长旺盛期的水热条件达到最佳,保证了红松宽轮的形成;上一年生长季末的降水有利于营养物质积累,促进了来年树木的生长,是形成红松宽轮的重要因素;在椴树红松林内处,生长季前的升温有利于红松打破休眠,及早进入生长季,也是影响红松生长的重要因子。
[1] Qian W H,Zhu Y F.Climate change in China from 1880 to 1998 and its impact on the environmental condition.Climatic Change,2001,50:419-444.
[2] Qian W,Qin A.Spatial-temporal characteristics of temperature variation in China.Meteorology and Atmospheric Physics,2006,93:1-16.
[3] Guo Q Y.Analysis of the intensity index and the change of East Asia wind.Acta Geographica Sinica,1983,38:207-217.
[4] Fritts H C.Tree rings and climate.London:Academic Press,1976.
[5] Chen Z J,Chen W,He X Y,Shao X M,Sun Y,Zhang C T,Liu T H.2007.Development of Chinese pine tree-ring width chronology in Fuling Mausolem,Shenyang of northeastern China.Journal of Beijing Forestry University,2007,29(4):100-109.
[6] Zheng Y H,Liang E Y,Zhu H F,Shao X M.Response of radial growth of Qilian juniper to climatic change under different Habitats.Journal of Beijing Forestry University,2008,30(3):7-12.
[7] Shao X M,Wu X D.Reconstruction of climate change on Changbai Mountain,Northeast China using tree-ring data.Quaternary Sciences,1997,3:76-85.
[8] Yu D P,Wang S Z,Tang L N,Dai L M,Wang Q L,Wang S X.Relationship between tree-ring chronology of Larix olgensis in Changbai Mountains and the climate change.Chinese Journal of Applied Ecology,2005,16(1):14-20.
[9] Yu D P,Wang Q L,Wang G G,Dai L M.Dendroclimatic response of Picea jezoensis along an altitudinal gradient in Changbai Mountains.Science in China:Series E,2006,49:150-159.
[10] Yu D P,Wang G G,Dai L M,Wang Q L.Dendroclimatic analysis of Betula ermanii forests at their upper limit of distribution in Changbai Mountain,Northeast China.Forest Ecology Management,2007,240:105-113.
[11] Gao L S,Zhang C Y,Zhao X H,Gadow K V.Gender-related climate response of radial growth in dioecious Fraxinus mandshurica Trees.Tree-Ring Research,2010,66(2):105-112.
[12] Chen L,Wu S H,Dai E F.Analysis of the tree-ring width chronologies of Pinus koraiensis and Larix olgensis on Changbai Mountains,Northeast China.Geographical Research,2011,30(6):1147-1155.
[13] Gao L S,Wang X M,Zhao X H.Response of Pinus koraiensis and Picea jezoensis var.komarovii to climate in the transition zone of Changbai Mountain,China.Chinese Journal of Plant Ecology,2011,35(1):27-34.
[14] Li G Q,Bai F,Sang W G.Different responses of radial growth to climate warming in Pinus koraiensis and Picea jezoensis var.komarovii at their upper elevational limits in Changbai Mountain,China.Chinese Journal of Plant Ecology,2011,35(5):500-511.
[15] Holmes R L.Computer-assisted quality control in tree-ring dating and measurement.Tree-Ring Bulletin,1983,43:69-78.
[16] Kendall M G.Rank Correlation Methods.London:Charles Griffin,1975.
[17] Korner C,Paulsen J.A world-wide study of high altitude treeline temperatures.Journal of Biogeography,2004,31:713-732.
[18] Biondi F,Waiul K.DENDROCLIM2002:a C++program for statistical calibration of climate signals in tree-ring chronologies.Computer and Geosciences,2004,30:303-311.
[19] Buckley B M,Wilson R J S,Kelly P E,Larson D W,Cook E R.Inferred summer precipitation for southern Ontario back to AD 610,as reconstructed from ring widths of Thuja occidentalis.Canadian Journal of Forest Research,2004,34:2541-255.
[20] Liang E Y,Shao X M,Hu Y X,Lin J X.Dendroclimatic evaluation of climate-growth relationships of Meyer spruce(Picea meyeri)on a sandy substrate in semi-arid grassland,North China.Trees,2001,15:230-235.
[21] Sang W G,Wang Y X,Su H X,Lu Z H.Response of tree ring width to rainfall gradient along the Tianshan Mountains of northwestern China.Chinese Science Bulletin,2007,52(21):2954-2962.
[22] Peng J F,Gou X H,Chen F H,Liu P X,Zhang Y,Fang K Y.Characteristics of ring-width chronologies of Picea crassifolia and their responses to climate at different elevations in the Anyemaqen Mountains.Acta Ecologica Sinica.2007,27(8):3268-3276.
[23] Mäkinen H,Nöjd P,Kahle H P,Neumann U,Tveite B,Mielikäinen K,Röhle H,Spiecker H.Large-scale climatic variability and radial increment variation of Picea abies(L.)Karst.in central and northern Europe.Trees,2003,17:173-184.
[24] Kujansuu J,Yasue K,Koike T,Abaimov A P,Kajimoto T,Takeda T,Tokumoto M,Matsuura Y.Climatic responses of tree-ring widths of Larix gmelinii on contrasting north-facing and south-facing slopes in central Siberia.Journal of Wood Science,2007,53:87-93.
[25] Shao X M.Radial growth of Huashan Pine and its response to climate.Journal of Chinese Geography,1994,4(3/4):88-102.
[26] Shao X M,Wu X D.Tree-ring chronologies for Pinus armandi Franch from Huashan,China.Acta Geographica Sinica,1994,49(2):174-181.
[27] Zhao Z J,Tan L Y,Kang W D,Liu Q J,Li J Q.Responses of Picea likiangensis radial growth to climate change in the Small Zhongdian area of Yunnan Province,Southwest China.Chinese Journal of Applied Ecology,2012,23(3):603-609.
[28] Yu D P,Wang Q W,Wang Y,Zhou W M,Ding H,Fang X M,Jiang S W,Dai L M.Climatic effects on radial growth of major tree species on Changbai Mountain.Annals of Forest Science,2011,68:921-933.
[29] Dittmar C,Zech W,Elling W.Growth variation of common beech(Fagus sylvatica L.)under different climatic and environmental conditions in Europe-a dendroecological study.Forest Ecology and Management,2003,173:63-78.
[30] Peng J F,Gou X H,Chen,F H,Li J B,Liu P X,Zhang Y.Altitudinal variability of climate-tree growth relationships along a consistent slope of Anyemaqen Mountains,northeastern Tibetan Plateau.Dendrochronologia,2008,26:87-96.
[31] Wilson R J S,Luckman B H.Tree-ring reconstruction of maximum and minimum temperatures and the diurnal temperature range in British Columbia,Canada.Dendrochronologia,2002,20:257-268.
[32] Frank D,Eeper J.Characterization and climate response patterns of a high-elevation,multi-species tree-ring network in the European Alps.Dendrochronologia,2005,22:107-121.
参考文献:
[3] 郭其蕴.东亚夏季风强度指数及其变化的分析.地理学报,1983,38(3):207-217.
[5] 陈振举,陈玮,何兴元,邵雪梅,孙雨,张春涛,付印东,刘铁红.沈阳福陵油松年轮度年表的建立.北京林业大学学报,2007,29(4):100-109.
[6] 郑永宏,梁尔源,朱海峰,邵雪梅.不同生境祁连圆柏径向生长对气候变化的响应.北京林业大学学报,2008,30(3):7-12.
[7] 邵雪梅,吴祥定.利用树轮资料重建长白山区过去气候变化.第四纪研究,1997,17(1):76-85.
[8] 于大炮,王顺忠,唐立娜,代力民,王庆礼,王绍先.长白山北坡落叶松年表及其与气候变化的关系.应用生态学报,2005,16(1):14-20.
[12] 陈力,吴绍洪,戴尔阜.长白山红松和落叶松树轮宽度年表特征.地理研究,2011,30(6):1147-1155.
[13] 高露双,王晓明,赵秀海.长白山过渡带红松和鱼鳞云杉径向生长对气候因子的响应.植物生态学报,2011,35(1):27-34.
[14] 李广起,白帆,桑卫国.长白山红松和鱼鳞云杉在分布上限的径向生长对气候变暖的不同响应.植物生态学报,2011,35(5):500-511.
[22] 彭剑峰,勾晓华,陈发虎,刘普幸,张永,方克艳.阿尼玛卿山地不同海拔青海云杉(Picea crassifolia)树轮生长特性及其对气候的响应.生态学报,2007,27(8):3268-3276.
[26] 邵雪梅,吴祥定.华山树木年轮年表的建立.地理学报,1994,49(2):174-181.
[27] 赵志江,谭留夷,康东伟,刘琪璟,李俊清.云南小中甸地区丽江云杉径向生长对气候变化的响应.应用生态学报,2012,23(3):603-609.
