海岸带生态系统健康评价中能质和生物多样性的差异
——以江苏海岸带为例
2013-12-19唐得昊邹欣庆刘兴健
唐得昊,邹欣庆,* ,刘兴健
(1.南京大学地理与海洋科学学院,南京 210093;2.南京大学海岸与海岛开发教育部重点实验室,南京 210093;3.中国科学院南海海洋研究所边缘海地质重点实验室,广州 510301)
生态系统健康评估是当今生态学领域的研究热点之一。生态系统具有自组织和调节功能,由于影响该系统的因子十分复杂,因而无法通过简单的实验揭示系统各组分之间的关系[1]。根据Bossel的研究,稳定性是生态系统发展演变的必要条件[2]。物种(特别是起“上行”或“下行”效应的决定性物种)的消失往往危及生态群落甚至整个系统的稳定性,是生态系统发展演变过程中比较大的扰动[3]。Mar Arthur和Elton在20世纪70年代提出的多样性——稳定性理论仍是指导生物多样性和生态系统保护的理论基础[4-5]。目前,群落组成与结构指标、物种多样性和功能多样性指标、生态系统净生产力、生态压力指标、生态能质和结构生态能质指标等,都被用于评估生态系统健康状况[6-9]。
生物多样性指数定量反映生物群落内物种多样性程度,是用来判断生物群落结构变化或生态系统稳定性的指标,其计算相对简单。由于生物多样性和生态系统健康的关系是学术界有争议的话题[10-11],不少学者在论证生物多样性与生态系统稳定性关系的基础上[12-14],建立生态系统健康评价模型[16-17]。在评判标准上,物种多样性丰富的生态系统对环境变化有更强的抵抗力和弹性。因此,生物多样性指数较高在一定程度上能促进生态系统健康[18]。
复杂生态系统表现特征是热力学上的开放性,组成物种多样,时空范围内高度异质性,以及系统组成物质之间的非线性相关[19]。Patten等认为隐藏在基因层面的适应性能反映生态系统面临环境变化时的适应能力,甚至是系统在时空格局下组织体的等级[20],因而生态系统发展演变状况不能仅从物种多样性方面测量。Jørgensen和Mejer等首次将生态能质的概念应用于生态学研究领域[21]。Odum认为它可以指示受干扰生态系统特征的改变[22]。生态能质指标不仅可以在时间尺度上度量生态系统的发展状况,还能从生态系统的结构及其所包含的微观信息上衡量系统的健康程度[23]。近几十年,能质指标常作为目标函数,用于解释生态系统结构的变化,描述系统的发展方向,表示扰动后系统的自组织能力,并从生态系统内部结构和稳定状态、系统复杂性和多样性、系统稳定性和弹性、系统活力以及生态系统物种组成的平衡上来说明整个生态系统的健康状况[24-25]。能质、结构能质和生态缓冲能力作为整体评估指标被用于生态系统健康评价,并且健康的生态系统应该具有较高的能质、结构能质和生态缓冲容量[26]。目前,运用能质指标评价生态系统健康状况所选取的研究对象大多是湖泊、河口[27-30],只有少数文献曾探讨过海岸泻湖和大陆架[31-33]
同样是评价生态系统健康状况,能质和生物多样性指标对生态健康状态的描述方式和表现形式是有差异的。本文以江苏省海岸带为例,通过比较能质和生物多样性评价结果,分析两者在海岸带生态系统健康评估体系中的功能差异及其原因,以期为生态系统健康评估指标的选取提供理论参考和依据。
1 研究区域概况
江苏省是我国沿海大省,介于东经 116°18'—121°57',北纬 30°45'—35°20'之间,海岸线全长 954km,分属连云港、盐城、南通3市的12个县和1个市区。该省海岸带地理位置优越,岸线北起苏鲁交界的绣针河口,南抵长江北口,并与上海市隔江相望,西以内陆为依托,东濒黄海,是沿海开放的前沿[34]。
2 研究方法
2.1 数据来源
本研究的相关数据依托于“908海洋专项调查项目”——江苏省海洋环境检测中心和江苏省海洋水产研究所2006年夏季(7—8月)的南黄海近海航测结果。航测数据包含各航次测得的不同级别(微型、小型、中型和大型)浮游生物、大型底栖生物、游泳动物(鱼、虾、蟹、头足类)等生物量信息,水环境物理化学信息以及潮间带生物调查结果。
从北到南沿江苏省海岸带,北部海州湾取5个站点,中部辐射沙脊群取4个站点,南部长江口以北区域取6个站点,共15个站点(图1),计算每个站点游泳动物和底栖生物的能质、结构能质和多样性指数,并分析它们之间的相关关系。
夏季,在江苏近岸海域采集到的大型底栖动物主要有多毛类(53种)、软体动物(41种)、甲壳动物(53种)、棘皮动物(14种)以及其他动物(9种)累计采集种类数为170种。
游泳动物主要是鱼类、虾类、蟹类和头足类。夏季航次鱼类的渔获种类主要为小黄鱼(Larimichthys polyactis),占鱼类总重的 25.75%,其次为鮸鱼(Miichthys miiuy),占 19.4%,银鲳(Pampus argenteus)列第三,占 18.7%。其余的还有刺鲳(Psenopsis anomala)、斑鰶(Konosirus punctatus)、赤鼻棱鳀(Thryssa kammalensis)、海鳗(Muraenesox cinereus)、皮氏叫姑鱼(Johnius belengerii)等;虾类中,口虾蛄(Oratosquilla oratoria)渔获量最大,占虾类比例的54.87%,其次为戴氏赤虾(Metapenaeopsis dalei),占28.03%,再次为细巧仿对虾(Parapenaeopsis tenella),占4.83%;蟹类主要品种为三疣梭子蟹(Portunus trituberculatus),占蟹类总量的 53.1%,红线黎明蟹(Matuta planipes,占 26.56%),以及日本蟳(Charybdis japonica,占 12.32%);头足类中,日本枪乌贼(Loligo japonica)占总量的73.74%,短蛸(Octopus ocellutus)占15.37%,长蛸(Octopus variabilis)、金乌贼(Sepia esculenta)等所占比重不大。
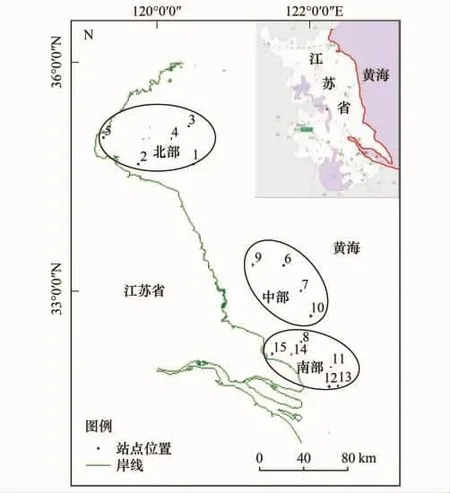
图1 采样点分布图Fig.1 Sampling points distribution
2.2 能质指标计算方法
能质是在一定环境条件下,系统为了达到同外界环境平衡时所需要做的最大总功。能质值(Ex)通过计算某一组分中的平均现存生物量(Ci)与其基因信息(βi)之间的乘积来获得[35]:

式中,Ex为生态能质(kJ/m3),R 为气体常数(kJ·g-1·K-1),T为绝对温度(K),Ci为生态系统中第 i种组分在现有状态的浓度(g/m3)。βi为第i种有机成分的权重转换因子。
结构能质(Exst)是系统中平均单位质量的能质,即能质同总生物量的比值[26],其计算公式如下:

式中,Exst为结构能质(kJ/g),Ct表示为系统中有机成分的总浓度或总生物量(g/m3)。
其中,βi值可根据储存在有机体基因内的信息[36]或种水平上的基因组尺度来计算[37]。本文根据Jørgensen的研究,βi权重因子计算公式如下:

通过该公式,Jørgensen在有机体能质计算一文中,详细给出了低到岩屑、病原体,高到智人(现代人)共45个不同级别和种类生物组分的 βi权重值[38]。
2.3 生物多样性指数计算方法
生物多样性的测度中,使用最广泛的是物种丰度,它最直观也最容易统计;其次是Shannon指数、Simpson指数以及Margalef指数,其他指数使用相对较少[39]。本文选取常用的生物多样性指数计算公式(表1)。

表1 多样性指数计算方法Table 1 The calculation method of diversity index
3 研究结果与分析
3.1 能质和结构能质的空间分布情况
在能质和结构能质计算过程中,权重因子 βi的确定方式多种多样,例如,通过营养群落、广系分类编码基因、物种分类水平基因等尺度进行估算,但是目前的研究表明,不同 βi值计算方法对能质指标作为生态指标的影响甚微[40-41]。因此,本研究针对采集样本的生物品种,利用已有文献对β权重的详细解译和计算[38],选取适当的权重值计算能质指标(表2)。
江苏海岸带能质值空间分布趋势是南部>北部>中部,而结构能质值空间分布趋势为北部>南部>中部(图2)。由于这3个区域生物量、环境因子差别较大,除了辐射沙脊群(中部)能质和结构能质值都偏小外,长江北口(南部)和海州湾区域(北部)差异较为明显。

表2 用于估算能值的权重转换因子(β)Table 2 The conversion factor weight(β)to estimate Exergy

图2 江苏海岸带能质和结构能质空间分布图Fig.2 The spatial distribution of Exergy and Structure Exergy of coastal areas in Jiangsu
江苏海岸带15个样本点南北位置、离岸远近以及测量环境的差异,都可能影响站点能质指标空间分布情况。对这15个调查站点能质和结构能质数据进行系统聚类分析,采用组间联接和平方Euclidean距离进行区间度量(图3)。
夏季江苏省海岸带健康情况可分为四类(表3):站点 1、2、7、9、10、13、15 为第 1 类(Ⅰ),该类能质和结构能质值都较低,显著水平α=0.05时,能质值和海水中总磷含量呈正相关关系,相关系数R2=0.604,这类生态系统生物量不大,且包含的信息量少,海水中总磷含量随能质值增高而增加,其中15号站点海水总磷含量为最大值(0.45),1、9号站点海水总磷含量为最小值(0.18),波动较大,说明系统组织性、有序化程度低,稳定性低,系统较脆弱,一旦系统受到破坏后使之恢复到热平衡状态所需的能量低;3、4、5、6为第2类(Ⅱ),该类能质值较低,结构能质值较高,能质值和海水中悬浮物含量相关,相关系数 R2=0.478(α=0.05),3、4 号站点位于海州湾东部,海水中悬浮物含量极少,5号站点位于海州湾近岸区,6号点位于辐射沙脊群北翼,海水中悬浮物含量极大。海水中悬浮物过大或过小,都会影响生态系统稳定性,加上Ⅱ类生态系统生物量很小,包含的信息量最少,系统不稳定;8、11、12为第3类(Ⅲ),该类能质值较高,结构能质值偏低,系统生物量很大,包含的信息量较大,水环境指标近于平均值,波动小,组织性、有序化程度较高,比较稳定;14为第4类(Ⅳ),该类有较高的能质和结构能质,系统稳定性强,组织性、有序化程度高,生物信息量大,且系统受破坏后使之恢复到热平衡状态所需的能量多,受外界干扰敏感度低。综其所述,以能质为目标函数时,Ⅳ类生态系统健康,Ⅲ类生态系统亚健康,Ⅰ、Ⅱ类生态系统存在问题。
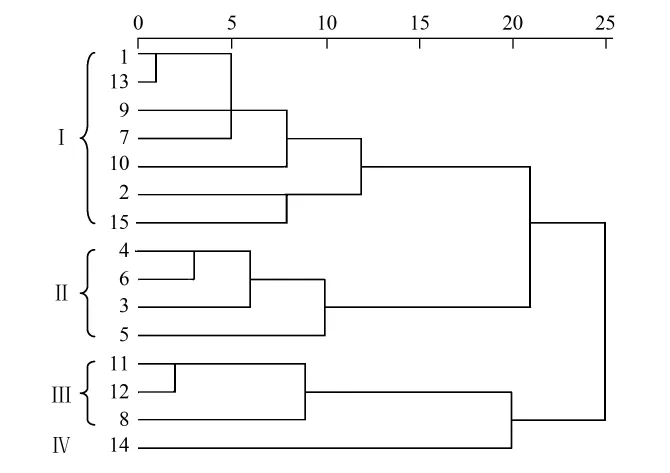
图3 江苏沿海站点生态能质的聚类分析Fig.3 Cluster analysis of Eco-Exergy of coastal areas in Jiangsu

表3 能质指标分类情况表Table 3 Exergy index classfication table
3.2 生物多样性指数空间分布情况
生物多样性指数测度一般可分为物种丰富度测度和异质性测度[42-43]。物种丰度简单指出群落中物种种数;Shannon指数作为异质性测度,可以测算物种多度的均匀程度。Shannon指数一般为1.5—3.5之间,只有当样本中出现大量物种时,才会有较大的值。May曾证明,若已知物种服从对数正态分布,要使Shannon指数>5.0,需要105种;Simpson指数侧重于样本多度较大的系统,通过它能得到物种多度分布的方差,群落组成越均匀,它的值就越大;Margalef指数是综合性指数,R.Margalef曾通过测得生态区域内生物多样性程度来判断区域污染程度[44]。
夏季,江苏沿海地区物种丰度值介于20—47之间,空间分布为北部>南部>中部;Shannon Wiener指数介于0.77—2.70之间,Simpson指数介于0.28—0.90之间,空间分布都为南部>中部>北部;Margalef指数介于2.50—5.50之间,南部略大于北部,中部较小(图4)。
物种丰度和Margalef指数最值出现地点一致,最大、最小值分别为8号点和6号点;以e为底的Shannon Wiener指数和Simpson指数最值出现地点一致,最大、最小值都分别出现在14号点和1号点。对生物多样性指数进行系统聚类分析(图5),得到4个类别(表4):站点2、9、11、12、14、15为第1类(Ⅰ'),该类生态系统Shannon Wiener指数和Simpson指数都相对较高,则物种丰富度和均匀度、优势度都比较高;与之相对的是第4类(Ⅳ',1号站点),ShannonWiener指数和Simpson指数都低;3、4、5、8为第2类(Ⅱ'),该类生态系统物种丰度和Margalef指数值高,物种丰富度大;与之相对的是第3 类(Ⅲ'),包括6、7、10、13 号点。因此,用生物多样性指数作为目标函数划分生态系统结果为:Ⅰ'、Ⅱ'类相对健康,Ⅲ'、Ⅳ'类生态系统存在问题。
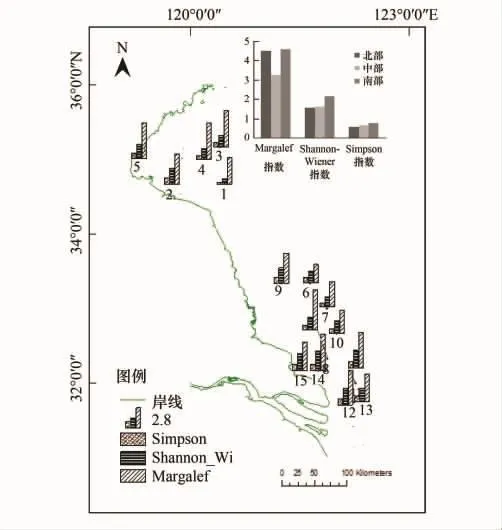
图4 江苏海岸带生物多样性空间分布图Fig.4 The spatial distribution of biodiversity of coastal areas in Jiangsu
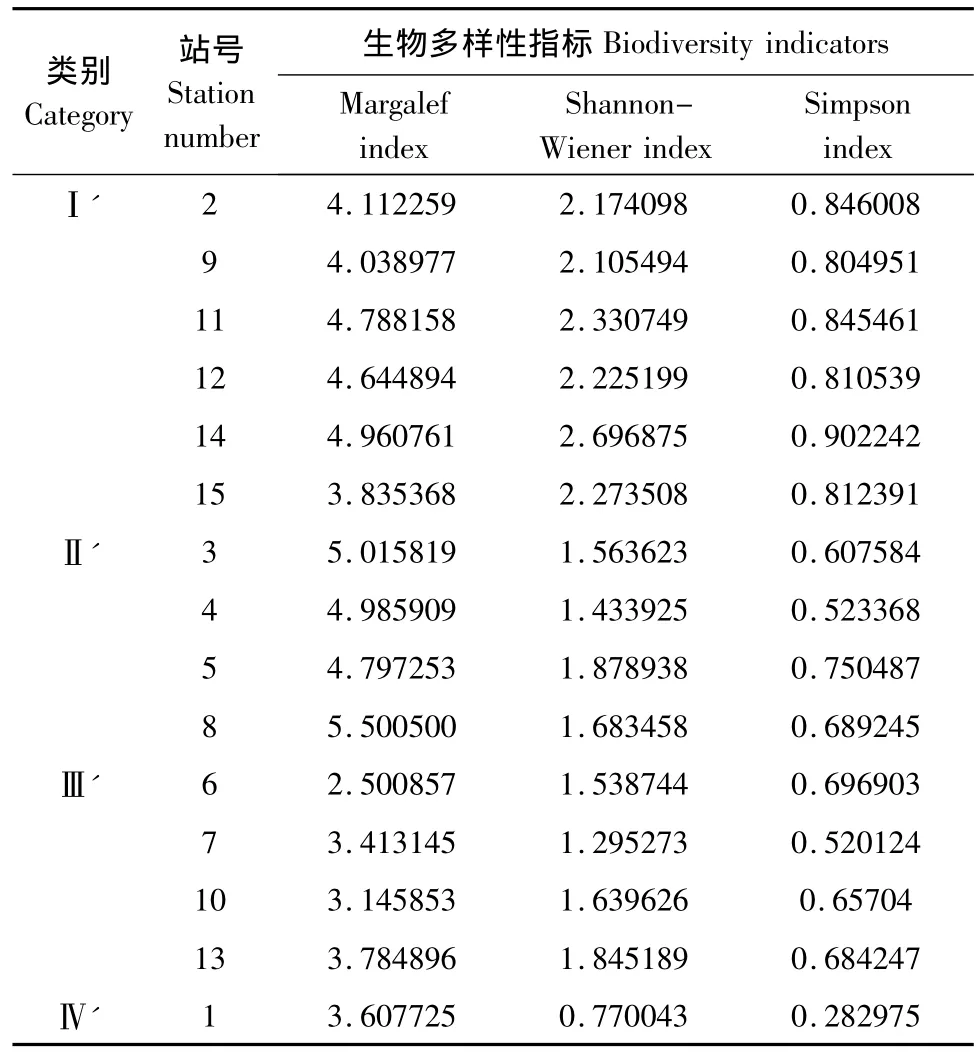
表4 生物多样性指标分类情况表Table 4 Biodiversity indicators classfication table
综上所述,不论以能质指标还是生物多样性指标作为目标函数,生态系统健康的站点有14号点,亚健康的站点为 8、11、12,不健康的站点为 1、6、7、10、13,其他站点(2、3、4、5、9、15)以生物多样性指标为目标函数时,被判定为健康的生态系统,而以能质指标为目标函数时,被判定为不健康的生态系统,该结果与两类指标测算偏重有关,前者主要从物种数量角度测量,后者侧重物种的基因组成及等级,数量大且种类多的生态系统不一定种群等级高,因此,最终判定这些站点的生态系统健康存在问题。
4 讨论与结论
4.1 能质与生物多样性指标存在一致性
能质值与生物多样性指标存在一定联系,分别将该两种指标作为目标函数,都可以分成四类,约2/3的站点健康状态评价结果一致。此外,能质和生物多样性最值(最大值、最小值)在一定程度上重合,说明能质与生物多样性指数能反映生态系统健康程度的总体趋势,并且两者反映的趋势一致。例如,14号点位于江苏海岸线南部近岸,同期温度较北部高,且海水中溶解态氮、总磷含量是所测点中的最大值,其环境有利于生物的生存和发展,能质和结构能质值都高,同时,该点也是Shannon Wiener指数和Simpson指数的最大值点;1号点位于海州湾东南侧远岸,受人类活动影响较小,总磷含量是所测点中的最小值,能质和结构能质值都低,同时是Shannon Wiener指数和Simpson指数的最小值点。
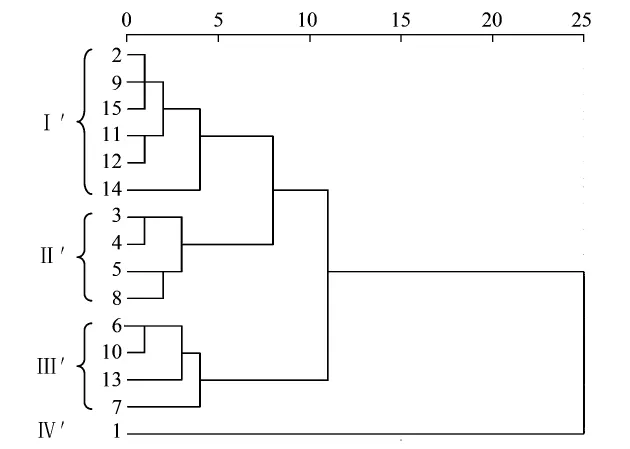
图5 江苏沿海站点生物多样性的聚类分析Fig.5 Cluster analysis of biodiversity of coastal areas in Jiangsu
4.2 能质和生物多样性指标在中低级生态系统中差异大
能质与生物多样性指标值排名前五的站点有80%的重合,其余排序中二者重合率低(表5),说明相对成熟的高级生态系统中,这两种指标能较一致地反映群落的健康状态,而在中低级生态系统中,能质与多样性指标差异较大。由于较高能质和结构能质的群落具有丰富的高级生物,且生态系统发育成熟,当群落或生态系统达到演替顶级以后,能维持其结构、功能长期保持在一个较高的水平[45],因此,这类生态系统处于一个比较平衡的状态。
相对而言,能质和生物多样性指标值较小的生态系统中,群落生物量偏小,生物等级较低,一旦受到外界干扰,波动较大,由某类生物数量激增或者外来物种入侵等情况引起的生物多样性指数和能质指标值变动的可能性大,因而这类演替中的群落处于非平衡状态。

表5 能质与生物多样性指标站点排名(自左向右降序排列)Table 5 Sample ranking of exergy and biodiversity(descending order from left to right)
4.3 能质与生物多样性指标关联程度低
能质指标中包含的种类级别以及营养类群信息是生物多样性指标所不能反映的。将能质指标与3个生物多样性指数做相关分析,发现能质与生物多样性指标的关联程度低。如图6所示,显著水平α=0.05时,能质与Margalef指数相关系数为R2=0.307,与Shannon Wiener指数相关系数R2=0.315,与Simpson指数相关系数 R2=0.243。
原因在于,能质值与物种等级和物种总量密切相关,生物多样性指数与组成生态系统各组分的物种等级和种类无关。已计算的能质值中,鱼类(β=499)等级最高,能质的权重β最大。据表6,14号点生物总量稍次于8号站点,且虾类(β=232)、蟹类(β=232)以及头足类(基本为软体动物,β=310)生物的能质排名靠后(第11),但由于14号站点鱼类总量大且种类丰富,能质值高,使该站点总能质值最大。8号点虽然生物总量最大,鱼类的能质值较14号点小,所以总能质小于14号点。因此,生物物种越高级,核苷酸数量多,所获权重大,反之亦反;生物量越大,能质值会增大,但总能质值不一定最高。同时,这也是将两类指标分别用于评判江苏省海岸带生态系统健康状态时,评价结果存在差异的原因。
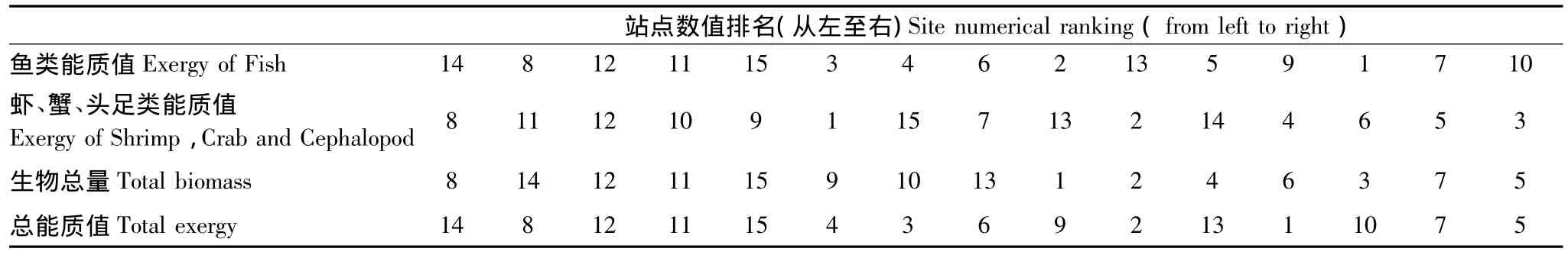
表6 调查站点数值排名表Table 6 Sorting table of sample stations
生物多样性测度与种群数量、种类生物体个数、分布均匀度等关联较大。以e为底的Shannon Wiener指数和Simpson指数的大小与群落中所有个体数量以及群落中第i个物种的个体数量直接相关。物种丰度和Margalef指数是群落中所有物种数目的函数,能在一定程度上说明生态系统物种丰度情况,但不能从物种种类和级别的角度阐述生态系统稳定情况。因此,能质和生物多样性指标侧重点不一,使得二者关联程度不高,评价结果不一致。

图6 能质与生物多样性指数相关度Fig.6 Correlation between Exergy and biodiversity
4.4 结论
综前所述,能质指标与生物多样性指数的差别如表7。
海岸带生态系统健康评价的过程中,指标体系的建立和评价方法仍然是核心。能质具有很好的热动力学理论基础,与系统论和信息论关系密切。通过能质指标能反映生态系统进化的水平,组织化水平和信息量,并定量生态系统的成熟程度。能质值越高,生态系统要达到热力学平衡状态需要从外界吸收的能量就越多,当生态系统的自组织结构功能发生变化,一定会在能质值上体现。生物多样性指标既能说明生态系统中物种生物量多样性和物种个体数量多样性,也能反映整个生态系统的结构层次和稳定情况。由于指标选取和指标衡量本身的局限性,在衡量生态系统健康状态时,能质指标可作为多样性指数等生态系统评价指标有益的、必要的补充。
[1] Jørgensen S E.Development of models able to account for changes in species composition.Ecological Modelling,1992,62(1/3):195-208.
[2] Bossel H.Real-structure process description as the basis of understanding ecosystems and their development.Ecological Modelling,1992,63(1/4):261-276.
[3] Pimm S L.The complexity and stability of ecosystems.Nature,1984,307(5949):321-326.
[4] MacArthur R.Fluctuations of animal populations,and a measure of community stability.Ecology,1955,36(3):533-536.
[5] Elton C S.The Ecology of Invasions by Animals and Plants.Chicago:University of Chicago Press,2000:143-153.
[6] Zhuang J R,Zhang Z X.Application of community structure-diversity index(CSDI)in forest health assessment in Beijing Badaling area.Chinese Agricultural Science Bulletin,2008,24(8):162-166.
[7] Chen Z Z,Qiu Y S,Jia X P,Huang Z R,Wang Y Z.Structure and function of Beibu gulf ecosystem based on Ecopath model.Journal of Fishery Sciences of China,2008,15(3):460-468.
[8] Wu T,Zhao D Z,Kang J C.Research progress on health evaluation of wetland ecosystem in valley-estuarine delta area.Marine Environmental Science,2010,29(2):286-292.
[9] Xu F,Yang Z F,Chen B,Zhao Y W.Ecosystem health assessment of the plant-dominated Baiyangdian Lake based on eco-exergy.Ecological Modelling,2011,222(1):201-209.
[10] May R M.Will a large complex system be stable?Nature,1972,238(5364):413-414.
[11] Walker B.Conserving biological diversity through ecosystem resilience.Conservation Biology,1995,9(4):747-752.
[12] Loreau M,Naeem S,Inchausti P,Bengtsson J,Grime J P,Hector A,Hooper D U,Huston M A,Raffaelli D,Schmid B,Tilman D,Wardle D A.Biodiversity and ecosystem functioning:current knowledge and future challenges.Science,2001,294(5543):804-808.
[13] Hooper D U,Chapin F S,Ewel J J,Hector A,Inchausti P,Lavorel S,Lawton J H,Lodge D M,Loreau M,Naeem S,Schmid B,Setälä H,Symstad A J,Vandermeer J,Wardle D A.Effects of biodiversity on ecosystem functioning:a consensus of current knowledge.Ecological Monographs,2005,75(1):3-35.
[14] Worm B,Barbier E B,Beaumont N,Duffy J E,Folke C,Halpern B S,Jackson J B C,Lotze H K,Micheli F,Palumbi S R,Sala E,Selkoe K A,Stachowicz J J,Watson R.Impacts of biodiversity loss on ocean ecosystem services.Science,2006,314(5800):787-790.
[15] Kimmins J P.Biodiversity and its relationship to ecosystem health and integrity.Forestry Chronicle,1997,73(2):229-232.
[16] Chen L,Wang X G.Biodiversity and forest ecosystem health:some key questions.Chinese Journal of Ecology,2008,27(5):816-820.
[17] Wang Y P,Wu H,Xu H C.Butterfly diversity and evaluation of forest eco-system health in important ecological areas of Zhejiang Province.Acta Ecologica Sinica,2008,28(11):5259-5269.
[18] Xu K Y,Ye W H.Ecosystem health and biodiversity.Ecologic Science,2002,21(3):279-283.
[19] Wu J G,Marceau D.Modelling complex ecological systems:an introduction.Ecological Modelling,2002,153(1/2):1-6.
[20] Constanza R,Jørgensen S E.Understanding and Solving Environmental Problems in the 21st Century.United Kingdom:Elsevier,2002:41-94.
[21] Jørgensen S E.Ecosystem theory,ecological buffer capacity,uncertainty and complexity.Ecological Modelling,1990,52(1/2):125-133.
[22] Odum E P.Trends expected in stressed ecosystems.Bioscience,1985,35(7):419-422.
[23] Sun P Y,Lu J J.Exergy theory:a new method of ecosystem study.Chinese Journal of Ecology,1997,16(5):32-37.
[24] Zhang J J,Jørgensen S E,Mahler H.Examination of structurally dynamic eutrophication model.Ecological Modelling,2004,173(4):313-333.
[25] Gurkan Z,Zhang J J,Jørgensen S E.Development of a structurally dynamic model for forecasting the effects of restoration of Lake Fure,Denmark.Ecological Modelling,2006,197(1/2):89-102.
[26] Li Q X,Feng J F,Zhu L.Applications of eco-exergy in ecological modeling and assessment of aquatic ecosystems.Chinese Journal of Ecology,2011,30(2):376-383.
[27] Fonseca J C,Pardal M A,Azeiteiro U M,Marques J C.Estimation of ecological exergy using weighing parameters determined from DNA contents of organisms — a case study.Hydrobiologia,2002,475-476(1):79-90.
[28] Xu F L,Zhao Y Z,Zhan W,Zhao S S,Dawson R W,Tao S.An ecosystem health index methodology(EHIM)for lake ecosystem health assessment.Ecological Modelling,2005,188(2/4):327-339.
[29] Zhang F J,Tong C F,Xie Z F,Liu W L,Gao W,Lu J J.The re-colonisation progress of intertidal benthic fauna community in the Changjiang Estuary.Acta Ecologica Sinica,2007,27(12):4944-4952.
[30] Zhai S J,Hu W P,Zhu Z C.Ecological impacts of water transfers on Lake Taihu from the Yangtze River,China.Ecological Engineering,2010,36(4):406-420.
[31] Salas F,Marcos C,Neto J M,Patrício J,Pérez-Ruzafa A,Marques J C.User-friendly guide for using benthic ecological indicators in coastal and marine quality assessment.Ocean and Coastal Management,2006,49(5/6):308-331.
[32] Austoni M,Giordani G,Viaroli P,Zaldívar J M.Application of specific exergy to macrophytes as an integrated index of environmental quality for coastal lagoons.Ecological Indicators,2007,7(2):229-238.
[33] Vassallo P,Fabiano M,Vezzulli L,Sandulli R,Marques J C,Jørgensen S E.Assessing the health of coastal marine ecosystems:a holistic approach based on sediment micro and meio-benthic measures.Ecological Indicators,2006,6(3):525-542.
[34] Ren M E.Comprehensive Investigation Report on Coastal Areas and Tidal Flat Resources in Jiangsu Province.Beijing:Maritime Press,1986:1-3.
[35] Austoni M,Giordani G,Viaroli P,Zaldivar J M.Application of specific exergy to macrophytes as an integrated index of environmental quality for coastal lagoons.Ecological Indicators,2007,7(2):229-238.
[36] Jørgensen S E.Application of exergy and specific exergy as ecological indicators of coastal areas.Aquatic Ecosystem Health and Management,2000,3(3):419-430.
[37] Fonseca J C,Marques J C,Paiva A A,Freitas A M,Madeira V M C,Jørgensend S E.Nuclear DNA in the determination of weighing factors to estimate exergy from organisms biomass.Ecological Modelling,2000,126(2/3):179-189.
[38] Jørgensen S E,Ladegaard N,Debeljak M,Marques J C.Calculations of exergy for organisms.Ecological modeling,2005,185((2/4):165-175.
[39] Sun J,Liu D Y.The application of diversity indices in marine phytoplankton studies.Acta Oceanologica Sinica,2004,26(1):62-75.
[40] Debeljak M.Applicability of genome size in exergy calculation.Ecological Modelling,2002,152(2/3):103-107.
[41] Libralato S,Torricelli P,Pranovi F.Exergy as ecosystem indicator:an application to the recovery process of marine benthic communities.Ecological Modelling,2006,192(3/4):571-585.
[42] McIntosh R P.An index of diversity and the relation of certain concepts to diversity.Ecology,1967,48(3):392-404.
[43] Good I J.The population frequencies of species and the estimation of population parameters.Biometrika,1953,40(3/4):237-264.
[44] Anne E M.Measuring Biological Diversity.Zhang F,translator.Beijing:Science Press,2011:39-72.
[45] Zhang J Y,Zhao H L.Review on the study of vegetation stability.Chinese Journal of Ecology,2003,22(4):42-48.
[46] Zhang J J,Gurkan Z,Jørgensen S E.Application of eco-exergy for assessment of ecosystem health and development of structurally dynamic models.Ecological Modeling,2010,221(4):693-702.
[47] Liu X H,Li X,Peng Z J.The discussion on calculation methods of biodiversity.Hebei Journal of Forestry and Orchard Research,2008,23(2):166-168.
参考文献:
[6] 庄健荣,张志翔.结构-多样性指数在八达岭地区森林健康评价中的应用.中国农学通报,2008,24(8):162-166.
[7] 陈作志,邱永松,贾晓平,黄梓荣,王跃中.基于Ecopath模型的北部湾生态系统结构和功能.中国水产科学,2008,15(3):460-468.
[8] 吴涛,赵冬至,康建成.流域-河口三角洲湿地生态系统健康评价研究进展.海洋环境科学,2010,29(2):286-292.
[16] 陈亮,王绪高.生物多样性与森林生态系统健康的几个关键问题.生态学杂志,2008,27(5):816-820.
[17] 王义平,吴鸿,徐华潮.浙江重点生态地区蝶类生物多样性及其森林生态系统健康评价.生态学报,2008,28(11):5259-5269.
[18] 许凯扬,叶万辉.生态系统健康与生物多样性.生态科学,2002,21(3):279-283.
[23] 孙平跃,陆健健.埃三级(Exergy)理论:生态系统研究的一种新方法.生态学杂志,1997,16(5):32-37.
[26] 李晴新,冯剑丰,朱琳.生态能质(eco-exergy)在水生生态系统建模和评价中的应用.生态学杂志,2011,30(2):376-383.
[29] 章飞军,童春富,谢志发,刘文亮,高伟,陆健健.长江口潮间带大型底栖动物群落演替.生态学报,2007,27(12):4944-4952.
[34] 任美锷.江苏省海岸带和海涂资源综合调查(报告).北京:海洋出版社,1986:1-3.
[39] 孙军,刘东艳.多样性指数在海洋浮游植物研究中的应用.海洋学报,2004,26(1):62-75.
[44] Anne E M.生物多样性测度.张峰,译.北京:科学出版社,2011:39-72.
[45] 张继义,赵哈林.植被(植物群落)稳定性研究评述.生态学杂志,2003,22(4):42-48.
[47] 刘晓红,李校,彭志杰.生物多样性计算方法的探讨.河北林果研究,2008,23(2):166-168.
