鄱阳湖国家级自然保护区东方白鹳(Ciconia boyciana)种群数量变化与气候的关系
2013-12-17缪泸君李言阔谢光勇袁芳凯
缪泸君,李言阔,李 佳,谢光勇,袁芳凯
江西师范大学 生命科学学院,江西 南昌 330026
气候条件是影响生物生存的重要环境因子。气候及其变化对种群数量波动的影响是鸟类学家关注的焦点之一。近年来的研究表明,气候变化会对鸟类的种群数量动态产生较强的影响(Both et al,2006;Crick,2004)。例如,极端气候事件可能会直接导致死亡率的迅速提高,甚至导致局部种群的灭绝(Schreiber et al,1984)。天气条件也可能通过改变食物资源的数量和质量影响成体存活率和繁殖成功率,进而影响鸟类的种群数量(Cormont et al,2012)。该影响具有一定的时滞性,即在生活史某一阶段的气候条件对种群的影响需经过一定的时间方才显现(Yalden & Pearce-Higgins,1997)。某些留鸟成体的存活率与非繁殖期的天气条件存在显著相关性(Jones et al,2003),越冬期的不利天气将导致迁徙鸟类繁殖成功率下降,第二年种群数量下降。譬如,大天鹅(Cygnus cygnus)在越冬地幼体的比例与上一年冬季的平均气温显著相关,冬季气温将影响后期的繁殖成功率(Nilsson,1979)。
大多数迁徙鸟类种群数量的年际变化主要受其繁殖地生存条件的影响,但也有研究发现某些迁徙鸟类的种群数量受其越冬地生存条件的影响,如水蒲苇莺(Acrocephalus schoenobaenus)、崖沙燕(Riparia riparia)及草鹭(Ardea purpurea)等(Peach et al,1991;Bryant et al,1995;Den Held,1981)。对于雁鸭类等水禽,其后期的繁殖成功率还受越冬地和迁徙停歇地食物条件等因素的影响(Newton,2004)。因此,迁徙鸟类种群动态制约因素研究需考虑繁殖地、越冬地及迁徙停歇地的环境条件。
东方白鹳(Ciconia boyciana)全球种群现存仅~3 000只个体,被 IUCN红皮书列为濒危物种(Birdlife International,2013),也是我国国家I级重点保护野生动物(China Wildlife Propagation Institution for Protection,1989)。东方白鹳繁殖地主要位于中国和俄罗斯交界的黑龙江和乌苏里江流域,越冬地为我国长江中、下游地区(Birdlife International,2013)。其中,鄱阳湖区是东方白鹳在长江中、下游地区的主要越冬地。近年来,该区越冬东方白鹳种群数量表现出较大波动(Li et al,2011)。本研究分析鄱阳湖国家级自然保护区(以下简称保护区)1985—2011年(27 a)越冬东方白鹳种群数量的年际变化与越冬地气候变化的相关性及时滞效应,以及基于越冬地气候条件的种群数量波动模型与种群波动趋势的拟合程度。
1 材料与方法
1.1 研究地区概况
鄱阳湖湿地是我国最大的淡水湖泊湿地,位于长江南岸的江西省北部(E115°49′~116°46′,N28°11′~29°51′)。鄱阳湖为季节性吞吐型湖泊,承接赣江、抚河、信江、饶河及修水五大河来水,并受长江来水影响,水位年变幅达9.79~15.36 m,4月进入汛期,呈湖相,7月达最高水位,11月进入枯水期,并持续至翌年3月,表现为典型的水陆交替湿地景观(Liu et al,2011)。气候特征属亚热带湿润季风型气候,热量丰富,雨量充沛,无霜期长,四季分明(Wang,2004)。1980—2011年鄱阳湖区年平均温度为18.0 ℃,并呈上升趋势,气温季节性变化明显,年平均降水量1 607 mm,年际变化大,无明显增减趋势(图1)。
鄱阳湖湿地已记录迁徙水鸟310种。其中,冬候鸟 155种,夏候鸟 107种,是白鹤(Grus leucogeranus)、白头鹤(G. monacha)、白枕鹤(G.vipio)、灰鹤(G.grus)、东方白鹳、小天鹅(Cygnus columbianus)等珍稀濒危鸟类的重要越冬地(Wu et al,2002)。
1.2 研究方法
保护区 1985—2011年东方白鹳越冬种群数量数据来源包括:(1)1985—2000年、2002—2006年以及 2009年保护区东方白鹳年度最大种群数量(Wu & Ji,2002;Li et al,2011);(2)2007、2008、2010及2011年保护区东方白鹳年度最大种群数量(Zhu et al,2012a,b)。(3)1998 年及 2001 年环鄱阳湖水鸟调查所记录的东方白鹳种群数量(Li et al,2013)。其中,历年的东方白鹳种群数量均为保护区越冬种群的最大记录值。根据候鸟到达和迁离鄱阳湖的时间,将东方白鹳的越冬期分为越冬初期(当年10月—11月)、越冬中期(当年12月—翌年1月)及越冬后期(翌年2月—3月)。气象数据引用《中国地面气候资料月值数据集》南昌监测站1975—2011年的月值资料。
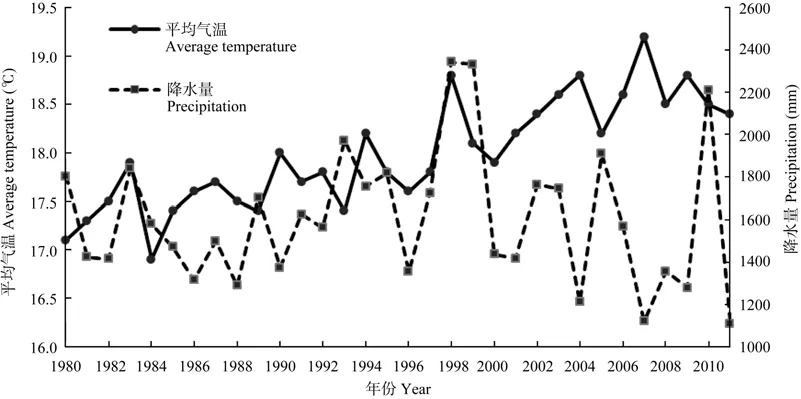
图1 鄱阳湖区1980—2011年平均气温与降水量变化(基于南昌气象站1980—2011年气象数据)Figure 1 Changes in annual average temperature and precipitation in the Poyang Lake from 1980 to 2011 (based on the data of Nanchang weather station)
以种群数量为因变量,年份为自变量,采用曲线回归法分析保护区 1985—2011年东方白鹳越冬种群数量的年际变化。利用 Kolmogorov-Smirnov test及Pearson相关分析检验种群数量动态与越冬期各月平均气温、平均最低与最高气温以及月降水量等气候变量的相关性。检验越冬地气候变量与其后 10年内东方白鹳种群数量的相关性以分析气候变量影响的时滞效应。同时,对东方白鹳种群数量动态相关性分析结果进行检验以消除极端值的影响。此外,以保护区东方白鹳种群数量为因变量,以与其显著相关的气候变量作为自变量,进行Stepwise多元线性回归分析,建立保护区东方白鹳种群数量动态预测模型,并使用VIF值进行共线性诊断(Yang & Zhang,2000;Yang et al,2012;Chatterjee & Price,1991)。数值统计以mean±SE表示,数据分析在Spss13.0中完成。
2 结 果
2.1 东方白鹳种群数量年际变化
1985—2011年,保护区东方白鹳平均越冬种群数量为(1 340±177)只,最小种群数量出现在1992年,为321只,最大种群数量出现在2010年,为3446只,年际波动较大(图2)。自1998年开始,越冬种群数量明显增加,1984—1997年,为(609±49)只,1998—2011 年,为(2 018±215)只,种群数量整体呈显著线性增长趋势(R2=0.735,F=69.414,df=26,P<0.01)。
2.2 东方白鹳种群数量与越冬地气候变量的关系
1985—2011年,越冬初期,保护区东方白鹳种群数量与越冬初期 10月份降水量(r=0.069,P=0.731,n=27)、平均气温(r=0.323,P=0.100,n=27)、平均最低气温(r=0.378,P=0.052,n=27)及平均最高气温(r=0.205,P=0.305,n=27)均不相关;与 11月份降水量(r=0.110,P=0.585,n=27)、平均最高气温(r=0.209,P=0.306,n=26)及平均气温(r=0.317,P=0.115,n=26)均不相关;与11月平均最低气温显著正相关(r=0.554,P=0.003,n=27)。
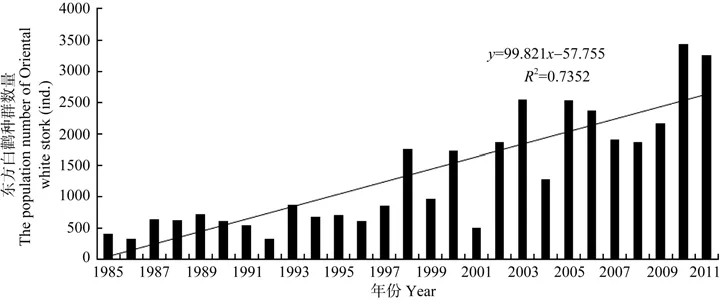
图2 鄱阳湖国家级自然保护区1985—2011年东方白鹳种群数量年际变化Figure 2 Annual population dynamics of the Oriental white stork in the PLNNR from 1985 to 2011
越冬中期,种群数量与12月份降水量(r=0.113,P=0.573,n=27)、平均气温(r=0.080,P=0.692,n=27)、平均最低气温(r=0.201,P=0.315,n=27)及平均最高气温(r=−0.059,P=0.771,n=27)均不相关,与 1 月份降水量(r=−0.0160,P=0.424, n=27)、平均气温(r=−0.230,P=0.249,n=27)、平均最低气温(r=−0.139,P=0.490,n=27)及平均最高气温(r=−0.303,P=0.124,n=27)均不相关。
越冬后期,种群数量与 2月份降水量(r=−0.031,P=0.878,n=27)、平均气温(r=0.192,P=0.337,n=27)、平均最低气温(r=0.195,P=0.329,n=27)及平均最高气温(r=0.191,P=0.339,n=27)均不相关,与3月份降水量(r=−0.171,P=0.394,n=27)、平均气温(r=0.270,P=0.173,n=27)、平均最低气温(r=0.210,P=0.293,n=27)及平均最高气温(r=0.306,P=0.120,n=27)均不相关。
2.3 气候对种群数量影响的时滞效应
考虑时滞效应,越冬初期,10月平均最低气温与一年后(r=0.461,P=0.016,n=27)及两年后(r=0.579,P=0.002,n=27)的东方白鹳种群数量均显著正相关;10月份平均气温与两年后(r=0.730,P=0.000,n=27)及 5年后的种群数量(r=0.511,P=0.006,n=27)均显著正相关;10月平均最高气温与两年后(r=0.777,P=0.000,n=27,图3)、四年后(r=0.612,P=0.001,n=27)及 5年后的种群数量(r=0.501,P=0.008,n=27)均显著正相关;11月平均气温(r=0.472,P=0.013,n=27)及 11月平均最低气温(r=0.463,P=0.015,n=27)均与四年后的种群数量显著正相关。
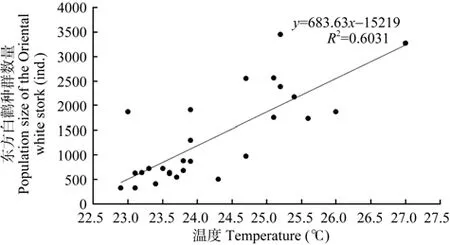
图3 鄱阳湖国家级自然保护区1985—2011年东方白鹳种群数量与越冬地2年前10月份平均最高气温相关性Figure 3 Correlations of the population size of the Oriental white stork in the PLNNR and the average maximum temperature of Oct. 2 years ago
越冬中期,12月气候变量与其后10年内东方白鹳种群数量动态变化无显著相关性,1月平均最低气温与5年后的种群数量显著正相关(r=0.427,P=0.026,n=27)。
越冬后期,2月份平均气温与两年后(r=0.471,P=0.013,n=27)、8年后(r=0.610,P=0.001,n=27)及 9 年后(r=0.557,P=0.003,n=27)的种群数量均显著正相关;2月份平均最高气温与两年后(r=0.449,p=0.019,n=27)、8 年后(r=0.580,P=0.002,n=27)及9年后(r=0.444,P=0.020,n=27)的种群数量均显著正相关;2月份平均最低气温与8年后(r=0.602,P=0.001,n=27)及 9 年后的种群数量(r=0.628,P=0.000,n=27)均显著正相关。3 月份平均气温与一年后(r=0.396,P=0.041,n=27)、两年后(r=0.483,P=0.026,n=27)、3年后(r=0.461,P=0.045,n=27)、4年后(r=0.705,P=0.000,n=27,图 4 )、6 年后(r=0.452,P=0.018,n=27),9 年后(r=0.441,P=0.021,n=27)及10年后的种群数量(r=0.405,P=0.036,n=27)均显著正相关。

图4 鄱阳湖国家级自然保护区1985—2011年东方白鹳种群数量与越冬地4年前3月份平均气温的相关性Figure 4 Correlations of the population size of the Orientalwhite stork in the PLNNR and the average temperature of Mar.4 years ago
3月份平均最低气温与两年后(r=0.428,P=0.026,n=27)、3年后(r=0.408,P=0.035,n=27)、4年后(r=0.673,P=0.000,n=27,图 5)、6年后(r=0.383,P=0.049,n=27)及 9年后的种群数量(r=0.427,P=0.026,n=27)均显著正相关。
3月份平均最高气温与两年后(r=0.516,P=0.006,n=27)、3 年后(r=0.479,P=0.011,n=27)、4年后(r=0.672,P=0.000,n=27,图6)及6年后的种群数量(r=0.462,P=0.015,n=27)均显著正相关。
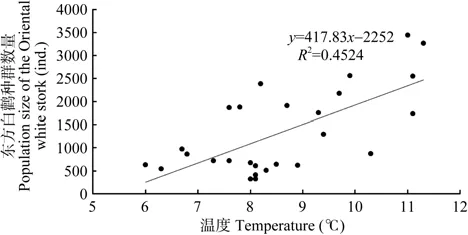
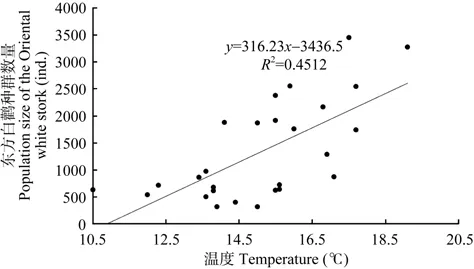
图6 鄱阳湖国家级自然保护区1985—2011年东方白鹳种群数量与越冬地4年前3月份平均最高气温相关性Figure 6 Correlations of the population size of the Oriental white stork in the PLNNR and the average maximum temperature of Mar. 4 years ago
2.4 基于气候变量的东方白鹳种群数量预测模型
将保护区 1985—2011年东方白鹳越冬种群数量作为因变量,与其存在显著相关性的气候变量作为自变量,进行Stepwise线性回归分析,结果表明,两年前的10月平均最高温度、4年前的10月最高温度、4年前的3月平均气温及两年前的3月平均最高气温为显著预测变量(表1)。其中,4个变量的 VIF值均<5,表明进入模型的变量不存在显著共线性。模型中的4个变量共同解释了保护区东方白鹳种群数量变化的79.2%(R2=0.792,F=20.901,df=26,P=0.000)。
回归模型为 Y=−15624+303.554X1+251.510X2+164.669X3+112.250X4,其中Y为保护区东方白鹳越冬种群数量,X1为两年前 10月平均最高气温,X2为4年前的10月平均最高气温,X3为四年前的3月平均气温,X4为两年前的3月平均最高气温。该模型较好地拟合了保护区东方白鹳种群数量变化趋势(图7)。
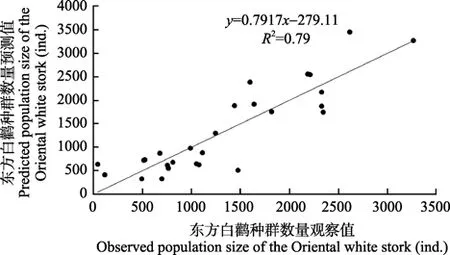
图7 基于越冬地气候条件的东方白鹳种群数量变化模型的预测值与观察值比较Figure 7 Comparisons of the predicted and observed population size of the Oriental white stork in the PLNNR
3 讨 论
在全球气候变暖的背景下,理解一个迁徙物种某一生活史阶段的气候条件如何对其后来的存活施加影响,已成为重要研究议题(McKellar et al,2012)。迁徙鸟类,尤其是长距离迁徙鸟类,其越冬地、迁徙停歇地和繁殖地气候条件差别极大,并均可能影响种群数量。东方白鹳主要在长江中、下游地区,尤其是鄱阳湖区越冬,越冬地相对固定。分析其越冬地气候变化对该种群的影响可以消除广泛分布区的影响,有利于揭示气候变化与种群波动的关系。本研究发现越冬地气温变量是东方白鹳种群数量波动的显著预测因子,提供了一个气候变化影响鸟类种群数量动态的例证。
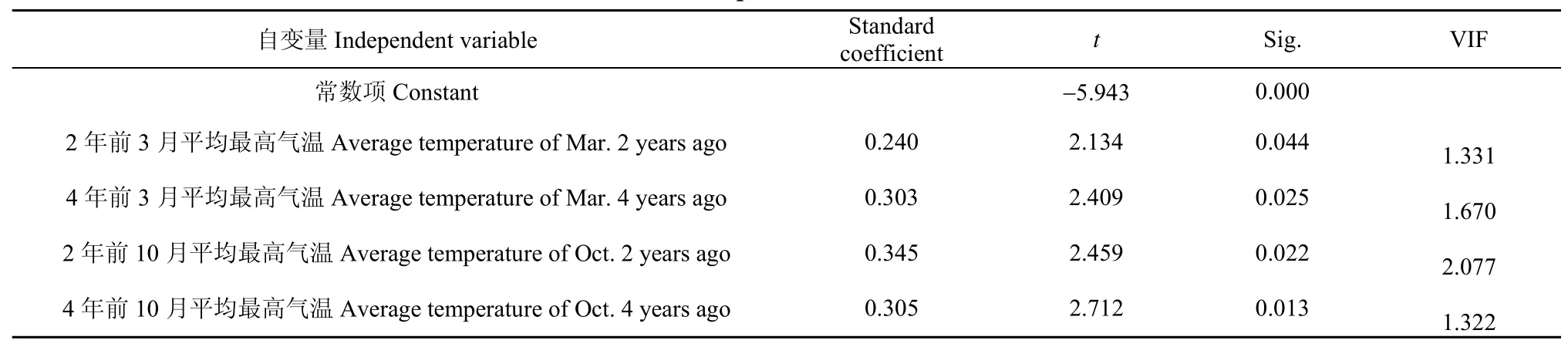
表 1 1985—2011年鄱阳湖国家级自然保护区东方白鹳种群数量与气候变量的线性回归分析Table 1 The stepwise linear regression analysis for dependent variable of the Oriental White Stork population size during1985−2011 and independent variables of climate
本研究发现东方白鹳种群数量与越冬当年 11月份的平均最低气温显著正相关,与越冬当年其他月份的气候变量均无显著相关性。一般来说,除非出现极端不利的气候事件,否则越冬当年的气候条件对东方白鹳种群数量通常不会产生很大的影响。东方白鹳一般于 10月底到达鄱阳湖,最早到达记录见于2001年和2005年的10月15日(Li et al,2011)。11月份是东方白鹳到达鄱阳湖的高峰,此时,最低气温显然不可能对东方白鹳个体产生直接作用。我们推测,由于不必像繁殖季节被限制在以巢址为中心的有限区域活动,越冬期鸟类相对自由,可以活跃移动,寻找食物资源丰富的地区。越冬初期的鄱阳湖低温可能会使东方白鹳在接下来的越冬期间往来迁飞于长江中、下游的众多湿地,寻找适宜的栖息地,增加对鄱阳湖以外区域的利用,导致在鄱阳湖区越冬个体数量的减少。卫星跟踪研究发现,东方白鹳个体在越冬期的飞行距离可达数百公里(Van den Bossche et al,2001),在安徽省的升金湖和江西省的鄱阳湖之间迁飞,越冬范围没有局限在单一的湿地内(Wu et al,2000)。
本研究首次发现越冬地气候条件对东方白鹳种群数量变化的影响存在显著的时滞效应,即越冬地气温对种群的影响至少在1年后开始变得明显,并与4年后种群数量的变化存在极显著相关性,越冬期气温的升高伴随着4年后种群数量的极显著增加。该时滞效应可能与气候对幼体的影响以及东方白鹳的性成熟年龄有关。近年来,有研究发现有东方白鹳个体 2岁龄达到性成熟(Jiang,2013),适宜的冬季气候条件更有利于幼鹳度过越冬期,使得更多的个体得以存活到繁殖年龄,并于2年后参加繁殖,导致2年后种群数量的增加。该时滞效应也见于气候对白鹤种群的影响。鄱阳湖冬季气候对白鹤种群数量的影响在3年后开始明显,可能与白鹤的性成熟年龄在3—5年有关(Li et al,2013)。
反之,越冬地恶劣的气候会导致几年后东方白鹳种群数量的减少。一方面,鄱阳湖水面结冰会对涉禽造成直接伤害,并可能造成个体数量的减少。如 2012年冬季,在对保护区巡护监测中曾发现一只东方白鹳被冰冻住双腿而受伤(Yu,2013)。我们在鄱阳湖区的鸟类调查过程中也曾两次发现死亡的东方白鹳个体。另一方面,冬季寒冷的天气会增加动物体温调节所需的能量(Hobbs,1989),而恶劣的冬季可能伴随着食物可获得性的下降,譬如,寒冷冬季雨后或雪后的结冰使东方白鹳无法取食,能量供求之间的不平衡会增加冬季死亡率,即使没有直接导致个体的死亡,也可能影响东方白鹳的能量积累,使其体况下降。
能量是影响迁徙鸟类种群动态的一个重要因子。若能量储存不足,一些个体将无法及时迁徙,从而留在越冬地(Van Dijk et al,1990)。在停歇地如果不能补充到充足的能量,个体尤其是幼体被捕食的概率会增大(Bijlsma & Jones,1990)。本研究发现,2年前的10月平均最高温度和3月平均最高气温、4年前的10月最高温度和3月平均气温是东方白鹳种群数量年际变化趋势的显著预测变量,共同解释了保护区东方白鹳种群数量变化的 79.2%。这两个月份是东方白鹳的越冬初期和越冬末期,是结束和重启长距离迁徙的阶段,可能是种群补充能量的关键时期,这两个时期越冬地恶劣的天气可能会导致个体无法获得充足的能量,不利于能量恢复和积累,从而对种群数量带来不利影响。
