Box-Behnken实验设计及响应面分析优化锰过氧化物酶培养基条件
2013-12-06赵玉萍陈晓旺沈鹏伟
赵玉萍,陈晓旺,沈鹏伟
(1.淮阴工学院生命科学与化学工程学院,江苏淮安 223001;2.江苏省生物质转化与过程集成工程实验室,江苏淮安223001)
随着世界化石能源危机和气候变化日益加剧,植物纤维作为可再生有机资源越来越受到重视,其主要成分为纤维素、木质素和半纤维素[1-2]。我国秸秆、稻壳资源丰富,但要实现该资源工业化利用还存在很多关键技术需解决,如利用有效的预处理技术打破木质素对纤维素的紧密结合,提高纤维素的降解效率等。随着生物技术的发展,现阶段在工业领域人们越来越热衷于木质素生物法的降解研究[2-3]。白腐菌是常用的木质纤维素降解微生物,其所含有的木质素降解酶系由木质素过氧化物酶(Lignin Peroxidase,简称LiP)、锰过氧化物酶(Manganese Peroxidase,简称MnP)和漆酶(Laccase,简称Lac)组成。其中,LiP和MnP同属于过氧化物酶,MnP和Lac又同属于酚氧化酶[4]。MnP也是一种糖蛋白,酶活性中心是由一个血红素基和一个Mn2+构成,还有两个起稳定结构作用的Ca2+。MnP表现出对Mn2+的绝对需要,对木质素的降解依赖Mn(Ⅱ)的存在[5],MnP先催化Mn2+转为Mn3+,Mn3+再去氧化大量的酚类底物。MnP氧化还原电位相对低的化合物,包括酚类木质素模式化合物的各种单酚和双酚,所以MnP被视为是一种酚氧化酶[6-7]。除了酚类化合物,MnP还能氧化胺类、染料,在纸浆、染料的脱色中起主要作用[8-10]。对于MnP的研究除了其催化降解机理外,大部分就是集中在提高酶的活性方面,采用不同的育种方法提高酶活、通过代谢调控酶量、优化培养条件提高酶活等[11-13]。为提高MnP酶活,本研究采用三水平部分因子分析法分析培养基中对酶活最重要的影响因素,然后采用Box-Behnken实验设计法及响应面分析法对培养基条件进一步优化,为MnP的进一步研究及工业化生产提供一定的依据。
1 材料与方法
1.1 材料与仪器
白腐真菌 由江苏省生物质转化与过程集成工程实验室筛选保藏;斜面培养基 PDA培养基;种子培养基 葡萄糖20g,硫酸铵3.5g,MgSO4·7H2O 0.5g,KH2PO41.0g,Na2HPO40.2g,MnSO40.035g,CuSO4·5H2O 0.007g,吐温-80 3mL,加蒸馏水至1000mL;初始发酵产酶培养基 麸皮10g,葡萄糖10g,酵母膏4.0g,酒石酸铵0.1g,MgSO4·7H2O 0.5g,KH2PO41.0g,Na2HPO40.2g,MnSO40.03g,CuSO4·5H2O 0.007g,吐温-80 3mL,加蒸馏水至1000mL。
自动立式电热压力蒸汽灭菌器 上海申安医疗器械厂;电热恒温培养箱 上海精宏实验设备有限公司;恒温摇床 江苏太仓市实验设备厂;离心机 上海安亭科学仪器厂;分光光度计 上海棱光技术有限公司。
1.2 实验方法
1.2.1 培养条件 250mL三角瓶中,种子培养基和产酶培养基装液量均为150mL,摇瓶中加入10颗玻璃珠。摇床条件均为:25℃、120r/min。种子培养时间为28h,接种量为10%。
1.2.2 酶活测定 粗酶液制备:取发酵液8000r/min离心10min,上清液即为粗酶液。
锰过氧化物酶活性测定:2mL反应液中,含有终浓度0.4mmol·L-1的愈创木酚,0.1mol·L-1H2O2,0.2mol·L-1MnSO4及50mmol·L-1的琥珀酸钠缓冲体系,酶液1mL,调节pH至4.5,加入0.5mL H2O2启动反应,30℃反应3min,测定反应液在波长465nm处单位时间吸光度的增加数,吸光度的增加是因愈创木酚氧化引起,愈创木酚氧化物呈茶黄色。一个酶活单位用每秒OD值增加0.1来表示[9,11,13]。
1.2.3 实验设计
1.2.3.1 三水平部分因子分析 根据前期实验的结果,选择了初始发酵产酶培养基中10个因子进行独立考察,利用Design-Expert 8.05软件设计三水平部分因子分析实验,加入4个中心点用于误差分析,初步确定主要的影响因子[14]。
根据初始发酵产酶培养基,设计各因素水平表,如表1所示。
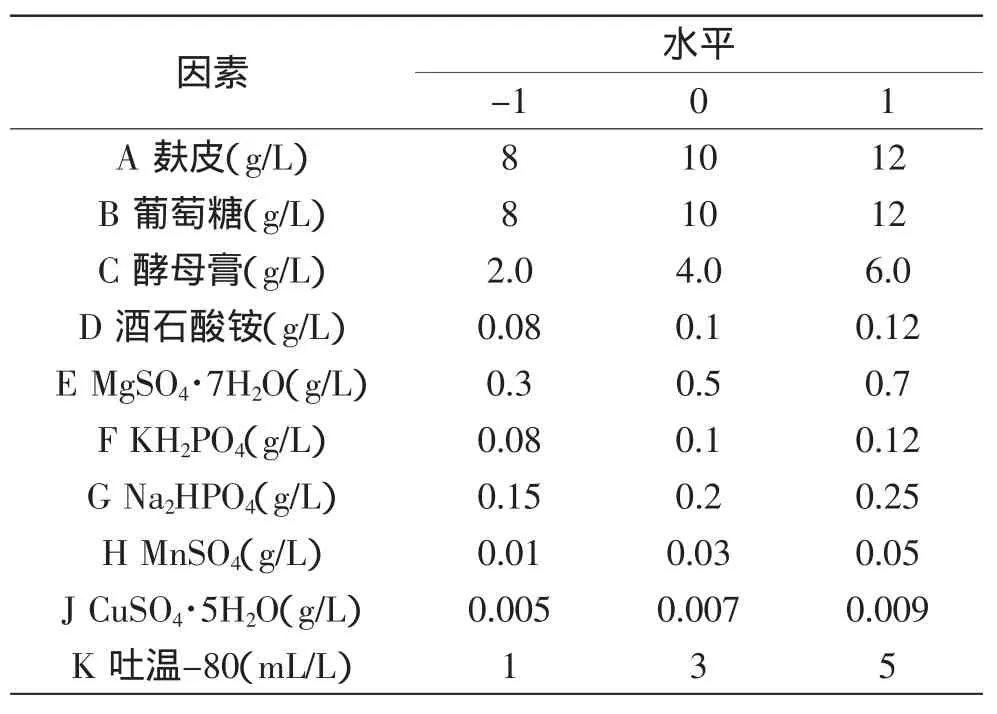
表1 部分因子分析实验各因素水平设计Table 1 Levels of the variables in 2-level factorial analysis
1.2.3.2 中心组合实验设计 筛选出影响锰过氧化物酶酶活的关键因子后,采用三因素三水平的中心组合设计法对关键因子进行优化。根据实验所得数据进行二次多项式回归拟合,预测锰过氧化物酶酶活的最大值[15]。三因素三水平的中心组合设计水平表见表2。
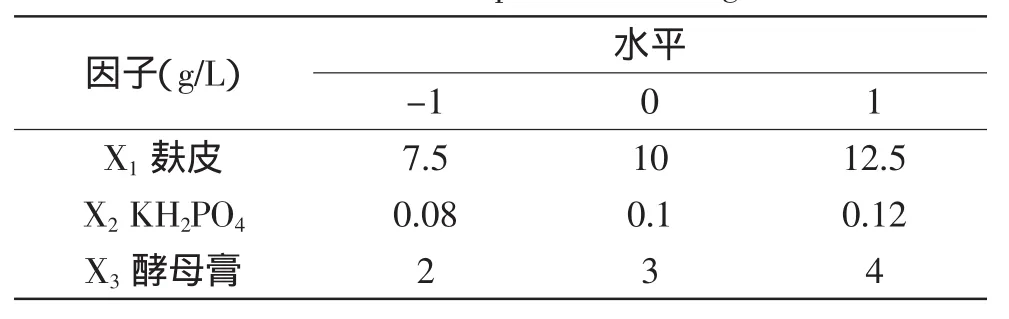
表2 Box-Behnken实验设计因子水平范围Table 2 Levels of the variables in the Box-Behnken experimental design
1.2.3.3 响应面模型的验证 以多元二次方程所预测的最佳培养基为最终发酵产酶培养基,进行发酵验证实验。
2 结果与讨论
2.1 初始锰过氧化物酶酶活测定结果
通过菌株的产酶发酵实验,根据锰过氧化物酶酶活测定方法,测定锰过氧化物酶酶活如图1所示。

图1 起始发酵培养基中锰过氧化物酶酶活曲线Fig.1 Manganese peroxidase activity curve of original enzyme production medium
由图1可以看出,菌株发酵至第3d开始有酶活,随后酶活逐渐上升,11d时酶活最高达2.62U/mL。随后酶活逐渐降低,23d时降至0.2U/mL,25d时几乎测不出酶活。呈现这样的变化主要是因为在最初培养基中营养物质主要供给菌体生长,因此酶活在3d前较低,随着菌体生长,在麸皮的诱导下,其产酶量逐渐上升,至11d达到最高峰,之后,菌体生长进入稳定期,随着营养物质的消耗,酶活开始下降。
2.2 三水平部分因子分析
按表1设计的因素水平表,根据软件,设计了20个实验点进行实验,测定结果如表3所示,对表3的数据进行方差分析,结果如表4所示。

表3 三水平部分因子分析实验设计与响应值表Table 3 Experimental design and response value in 2-level factorial analysis
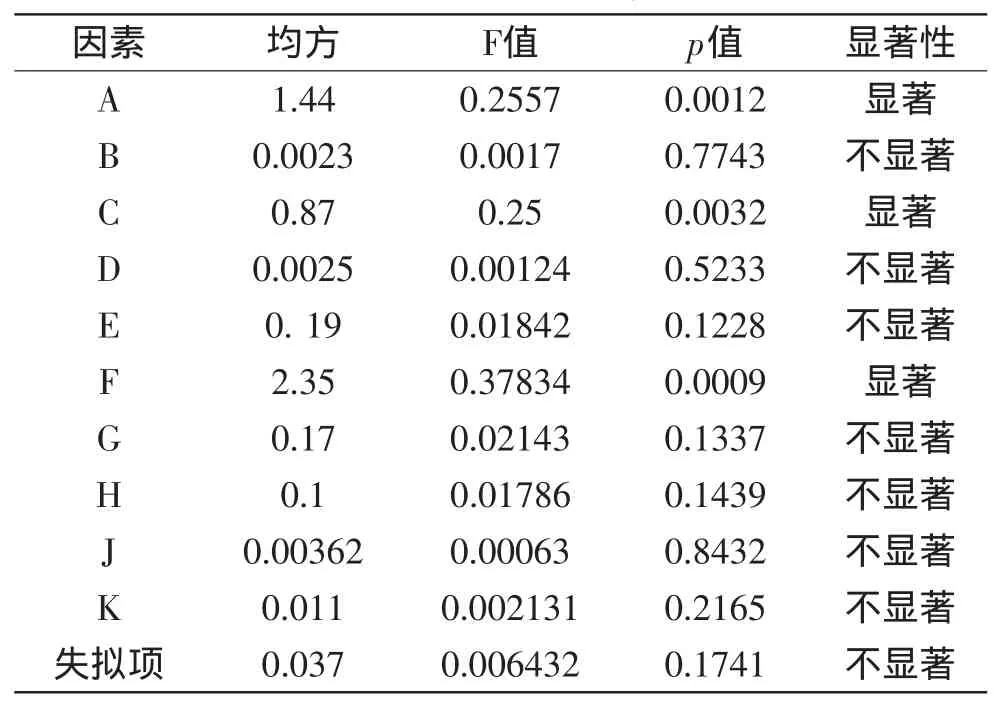
表4 三水平部分因子分析实验显著性结果Table 4 Test results of significance for 3-level factorial analysis
由表3和表4可知,显著影响因子是A(麸皮)、C(酵母膏)、F(KH2PO4)。因此选择以上三个因子进行以下实验,以进一步筛选最优添加量。
2.3 显著影响因子的Box-Behnken设计实验优化
根据Box-Benhnken的中心组合实验设计原理,进行三因素三水平的响应面分析实验,因子水平表如表2所示,实验共有17组,即17个实验点,其中析因点12个,另外5个为零点,为减少误差,零点实验重复5次。实验结果如表5所示。

表5 响应面实验结果Table 5 The result of Box-Behnken experiment
2.4 锰过氧化物酶的响应面结果与分析
利用Design-Expert 8.05软件对表5中的实验数据进行方差分析和回归拟合,以发酵液中锰过氧化物酶活为响应值Y,其结果如表6所示。
回归方程中各变量对响应值影响的显著性由F检验来判定,p值越小,则相应变量的显著程度越高。从表6的二次多项模型和方差分析结果看出,一次项系数Xl、X3,二次项系数X12、X32的p值均小于0.05,说明这些因素对菌株发酵产锰过氧化物酶酶活影响显著。而一次项系数X2,二次项系数X22,交互项X1X2、X1X3、X2X3的p值均大于0.05,说明三因素的交互项交互作用不显著,且以上5项对变异菌株发酵生产锰过氧化物酶酶活影响不显著。经过对实验数据的回归分析,得到二次多元回归方程:
Y=4.04+0.10X1+0.032X2+0.050X3+0.035X1X2-0.040X1X3-0.050X2X3-0.15X12-0.065X22-0.11X32。

表6 响应面实验方差分析结果Table 6 The significance analysis of Box-Behnken experiment
从表6中可以看出,复相关系数R2=92.89%,说明响应值的变化有92.89%来源于所选变量。失拟项p值为0.9941(>0.05),表明失拟不显著,因此该回归方程可以较好地描述各因素与响应值之间的真实关系,可以利用该回归方程对本实验产木质素酶降解菌株发酵产酶进行分析和预测。
根据响应面回归方程可以利用Design-Expert 8.05软件绘制响应曲面图,见图2~图4。每个响应面代表了当一个变量处于最佳水平时,另外两个独立变量之间的相互作用。
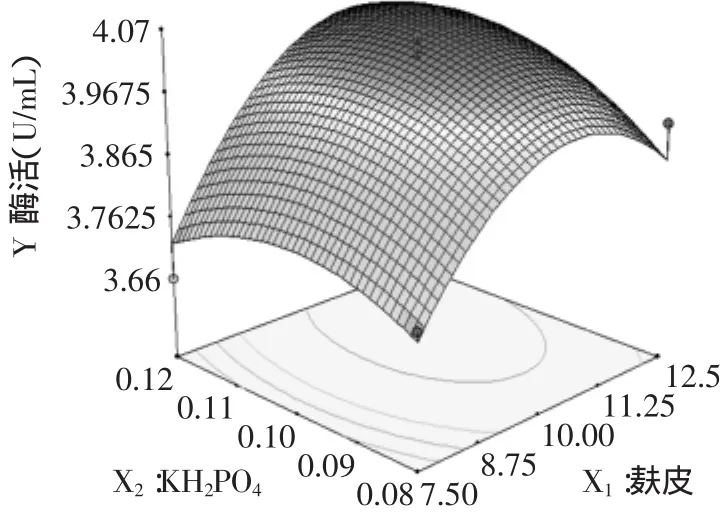
图2 麸皮与KH2PO4对锰过氧化物酶酶活的响应面曲面图Fig.2 Response surface plot of wheat bran and KH2PO4 on Mnp enzyme activity

图3 麸皮和酵母膏对锰过氧化物酶表达的响应面Fig.3 Response surface plot of wheat bran and yeast extract on Mnp enzyme activity

图4 酵母膏和KH2PO4对锰过氧化物酶表达的响应面Fig.4 Response surface plot of yeast extract and KH2PO4 on Mnp enzyme activity
每个响应面分别代表着两个独立变量之间的相互作用,此时第三个变量保持最佳水平。由图2~4可以看出,麸皮、酵母膏与锰过氧化物酶产量存在显著的相关性。当KH2PO4和酵母膏维持固定在最佳水平时,麸皮的添加量由7.5g/L升至10.75g/L时,锰过氧化酶产量随之升高,当超过10.75g/L时,锰过氧化物酶产量开始下降。
根据Design-Expert软件分析和预测,各主要因素麸皮、酵母膏和KH2PO4的优化值分别为10.75、3.37、0.095g/L,此条件下的酶活最佳值为4.06U/mL。
2.5 菌株发酵最佳条件的确定及其检验
为检验模型预测值与实际实验值之间的相关性,即检验响应面优化模型的可靠性,对菌株在预测的最优发酵条件下产酶能力进行实验验证。实验中麸皮、酵母膏和KH2PO4的优化值分别为10.75、3.37、0.095g/L,三组平行实验,测得其发酵后的Mnp酶活分别为4.14、4.23、4.07U/mL,平均值为4.15U/mL,与预测值十分接近。优化后的酶活与优化前相比,Mnp酶活提高了58.4%。
3 结论
综上所述,采用白腐真菌为出发菌株,根据Design-Expert 8.05软件设计的三水平部分因子分析初始发酵产酶培养基因子,确定了麸皮、酵母膏、KH2PO4为产Mnp的显著影响因子,根据Box-Benhnken中心组合实验设计及三因素三水平的响应面分析,通过二次多项回归模型进行方差分析和回归拟合,预测了最佳产酶培养基条件为:麸皮10.75g/L、酵母膏3.37g/L、KH2PO40.095g/L,最大Mnp酶活预测值为4.06U/mL。验证实验中Mnp酶活为4.15U/mL,与预测值十分接近,比优化前酶活提高了58.4%,证明了该实验方法的稳定性。
[1]陈洪章.纤维素生物技术[M].北京:化学工业出版社,2004:1-7.
[2]白娜,顿宝庆,宋立立,等.高活力锰过氧化物酶菌株的选育及其酶学性质初探[J].西北农林科技大学学报:自然科学版,2010,38(11):139-156.
[3]Mohammad J, Taherzadeh, Keikhosro K.Pretreatment of lignocellulosic wastes to improve ethanol and biogas production:A review[J].International Journal of Molecule Science,2008(9):162-165.
[4]黄茜,黄凤洪,江木兰,等.木质素降解菌的筛选及混合菌发酵降解秸秆的研究[J].中国生物工程杂志,2008,28(2):66-70.
[5]Monteiro L,Mariano RLR,Souto-Maior AM.Antagonism of Bacillus spp Against Xanthomonas campestris pv.Campestris[J].Brazilian Archives of Biology Technology,2005,48(1):23-39.
[6]Dehghan-Noude G,Housaindokht M,Bazzaz BS.Isolation,characterization,and investigation of surface and hemolytic activities of a lipopeptide biosurfactant produced by Bacillus subtilis ATCC 6633[J].Journal of Microbiology,2005,43(3):272-276.
[7]Chiang ML, Chou CC.Expression of superoxide dismutase,catalase and thermostable direct hemolysin by and growth in the presence of various nitrogen and carbon sources of heatshocked and ethanol-shocked Vibrio parahaemolyticus[J].International Journal of Food Microbiology,2008,121(3):268-274.
[8]阮久莉,王勐,毛亮,等.白腐菌锰过氧化物酶对2,2,4,4-四溴联苯醚的降解[J].环境科学与技术,2012,35(1):20-24.
[9]Kim YM,Nam IH,Murugesan K,et al.Biodegradation of diphenyletherand transformation ofselected brominated congeners by Sphingomonas sp.pH-07[J].Applied Microbiology and Biotechnology,2007,77(1):187-194.
[10]Chigu NL,Hirosue S,Nakamura C,et al.Cytochrome P450 Monoxygenases involved in anthracene metabolism by the whiterot basidiomycete Phanerochaete chrysosporium[J].Applied Microbiology and Biotechnology,2010,87(5):1907-1916.
[11]周德明,冯友仁,梁帅.木质素过氧化物酶和锰过氧化物酶高产菌株的诱变育种[J].中南林业科技大学学报,2009,29(5):115-118.
[12]白娜,顿宝庆,宋立立.高活力锰过氧化物酶菌株的选育及其酶学性质初探[J].西北农林科技大学学报,2010,38(11):139-145.
[13]Bonname P,Jeffries TW.Mn(Ⅱ)regulation of lignin peroxidases and manganese dependent peroxidases from ligindegrading white rot fungi[J].Applied and Environment Microbiology,1990,56:210-217.
[14]于平,陈益润.响应面法优化芽孢杆菌发酵生产植酸酶[J].中国粮油学报,2011,26(3):86-90.
[15]刘玲玲,陈钧.响应面法优化麦麸发酵产植酸酶条件的研究[J].中国粮油学报,2009,24(10):112-115.
