浙江中部地区公益林的群落结构
2013-12-05钱逸凡韩冰园伊力塔余树全郑超超
钱逸凡 ,韩冰园,伊力塔 ,张 超,余树全 ,郑超超
(1.浙江农林大学 林业与生物技术学院,浙江 临安 311300;2.国家林业局 华东林业调查规划设计院,浙江 杭州 310019;3.敦化林业有限公司,吉林 敦化 133700;4.浙江农林大学 亚热带森林培育国家重点实验室培育基地,浙江 临安 311300)
森林群落结构是群落生态学研究的基础[1],是指群落中的所有种类及其个体在时间和空间上的配置状况[2-4]。种群是群落的组成部分,其结构不仅对群落结构有直接影响,而且能客观地体现群落的发展趋势[5-8]。因而种群结构的研究对阐明群落的形成及其稳定性与演替规律等都具有重要意义[8-9]。亚热带地区的森林群落有其独特特点,群落类型多样、物种丰富、生产力高,一直是国内外研究的热点[10-13]。由于人类的过度利用,浙江省典型的亚热带森林群落遭到了巨大的破坏,带来了许多生态问题。为此,浙江省于2001年起全面启动实施 “200×104hm2生态公益林”计划,随后关于浙江省公益林的研究也逐渐增多[14-16],但由于浙江省的地貌为 “七山一水二分田”,立地类型多样、生境复杂,在省级尺度上的研究普遍存在样本少,缺乏系统性等问题。尤其是浙江省中部地区承接南北、区域面积大,且原始植被破坏早而剧烈,但目前尚无针对该区域公益林群落的专项研究。因此,在分析浙江省中部主要优势种群的基础上,探求公益林群落的组成和结构特征,对指导该地区植被恢复和公益林的经营管理具有重要的科学和现实意义。
1 研究区概况
浙江省位于中国长江中下游东南沿海地区,长江三角洲南翼,27°06′~31°31′N,118°01′~123°10′E。浙江省中部面积广阔,包括金华市的全境以及绍兴市、衢州市、台州市和丽水市的绝大部分地区。该地区开发历史悠久,经历了几千年人类活动和干扰,原生森林植被大多已破坏无遗,特别是近百年来几乎完全毁坏。20世纪50年代经济快速发展,人为活动频繁,环境再一次被破坏。20世纪90年代开始重视环境保护,森林恢复迅速,其公益林面积达165.46万hm2,占浙江省总面积 260.70万hm2的63.47%,是浙江省的生态建设的重点区域(表1)。在林龄结构上,浙江中部地区公益林相比全省的平均水平,幼龄林的比例更低为26.58%(图1)。在公益林群落的结构上,中部地区仍以针叶林为主,松林、杉木Cunninghamia lanceolata林比例分别为37.58%和15.29%,高于全省整体水平;此外,其阔叶林比例偏小,为24.65%,小于全省的28.48%,而针阔混交林具有较高的比例,为14.76%。

表1 浙江中部公益林面积统计Table 1 Area central statistics of public welfare forest in the central Zhejiang
2 研究方法
2.1 样地的选取与设置
2.1.1 样地的选取 由于浙江中部面积广大,要使分布样地均匀困难较大。本研究根据浙江省行政区划,选取可代表中部5市平均质量的县市进行重点观测,设置包括嵊州市(绍兴地区)、东阳市(台州地区)、仙居县(台州地区)、缙云县(丽水地区)、江山市(衢州地区)在内的公益林固定监测小班。在获取公益林二类资源清查数据的基础上,采用两阶抽样法。第1阶以各县公益林小班为抽样的基本单元,以全部公益林的小班为总抽样总体,按分层抽样抽取监测小班[14],共设监测小班517个,其中嵊州市96个,东阳市121个,仙居县103个,缙云县117个,江山市80个;第2阶在全面踏查小班的基础上,对每个抽样小班设置1个典型样地。
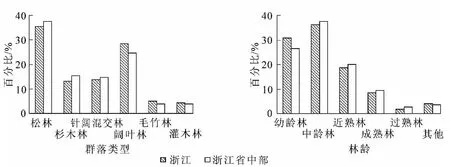
图1 公益林群落类型与林龄分布Figure 1 Community types and stand age of public welfare forest
2.1.2 样地的设置 在对固定监测小班全面踏查的基础上,选取典型地段内设置面积为20 m×20 m的固定样地,详细记录样地基本信息,包括经纬度、海拔、坡度、坡向、坡位、郁闭度、土壤等环境因子;样地内乔木层(胸径大于5.0 cm)采用每木调查(包括测定树高、胸径、冠幅和枝下高等),同时在每块样地对角线上均匀设置3个2 m×2 m的灌草固定小样方(图2),详细记录灌木种类、株数、盖度、高度以及草本种类、株数、盖度等指标,所有调查均在2010-2011年完成。

图2 样方设置示意图Figure 2 Setting and investigating in sampling site
2.2 数据处理
2.2.1 重要值 在群落的分析中,重要值的数值大小可作为群落中植物种优势度的一个度量标志,并可以体现群落中每种植物的相对重要性及植物的最适生境,其计算公式为:①乔木层重要值(%)=(相对高度+相对显著度+相对多度)/3。其中,相对高度(%)=(某个种的高度/所有种个体高度之和)×100;相对显著度(%)=(某个种的基径断面积/所有种的基径断面积之和)×100;相对多度(%)=(某个种的株数/所有种的总株数)×100。②灌木层、草本层重要值(%)=(相对盖度+相对多度)/2。其中:相对盖度(%)=(某个种的盖度/所有种的总盖度)×100;相对多度(%)=(某个种的株数/所有种的总株数)×100。
2.2.2 静态生命表 对树木种群年龄结构的研究多采用空间代替时间的方法,即以胸径级代替年龄进行分析[17-19]。对于亚热带树种,目前尚无统一的径级结构划分标准,常以 2~5 cm 划分为一个径级[3,20-21],因此,本研究按5 cm为一个径级统计各径级内的株数,编制主要优势种群静态生命表。静态生命表包括如下参数:nx,在x期开始时的存活数;lx,x期开始时的存活率;dx,从x到x+1的死亡数;qx,从x到x+1的死亡率;Lx,从x到x+1期的平均存活数,即Lx=(nx+nx+1)/2;Tx,进入x期的全部个体在进入x期以后的存活个体总年数,即Tx=∑Lx;ex,x开始时的生命期望或平均余年;Kx,环境阻力或消失率,表示种群从x期到x+1期间受到的阻力,Kx=ln lx-ln lx+1。
3 结果与分析
3.1 公益林的组成
将样地分成浙江省常见的松林、杉木林、针阔混交林、阔叶林、毛竹林和灌木林六大类型[14-15]。本次调查乔木层树种共52科103属263种,灌木层植物59科125属449种,草本层植物36科56属286种;群落物种数依次为针阔混交林>阔叶林>松林>灌木林>毛竹林>杉木林(表2)。多为人工构建的杉木林虽然在乔木层密度、平均胸径和平均树高上占据优势,但是植物种单一,群落结构相对不稳定,物种保育能力也往往不高;而松林多为天然次生林,乔木密度低,林下光照条件好,易于物种的入侵和定居,因此灌木层物种丰富,为向针阔混交林演替储存了物质基础;对于阔叶林而言,由于建国初期的过度利用,其典型的群落已所剩无几,故在发育上多为天然次生而来,目前在平均胸径和平均树高上均不占优势。

表2 浙江中部地区各公益林群落基本情况Table 2 Basic information of public welfare forest community
对本次调查中株数占样地总株数1%以上的植物中进行重要值分析(表3),乔木层中,马尾松Pinus massoniana(35.97)为最高;其次为杉木(15.64)和毛竹 Phyllostachys edulis(12.40);木荷 Schima superba,青冈Cyclobalanopsis glauca,黄山松Pinus taiwanensis和枫香Liquidambar formosana等也都有较高的重要值。灌木层中,檵木Loropetalum chinensis(11.84)重要值最高,其次为映山红 Rhododendron simsii(8.86);此外,柃木Eurya japonica,枫香,箬竹 Indocalamus tessellatus,白栎 Quercus fabri等也有较高的重要值。草本层中,大叶狼衣 Osmunda japonica(11.84)具有最高的重要值,其次为香附子Cyperus rotundus(15.23)和欧洲蕨 Pteridium aquilinum(8.31),芒 Miscanthus sinensis(4.19),千金子 Leptochloa chinensis(4.15)等。通过比较不同公益林群落植物种的重要值,发现各个群落在不同层次中的植物种类与组成,普遍具有建群树种少,树种构成相对集中等特点,在灌木层和草本层中尤为明显,各群落灌木层以檵木、柃木、杜鹃等小乔木为主,而草本层重要值最大均为强阳性的大叶狼衣。各群落乔木层的优势种差异较大,松林和针阔混交林乔木层中的重要值最大的均为马尾松,分别为83.07和39.80,杉木林则以杉木(73.39)重要值最大,阔叶林以木荷(19.21)和青冈(10.73)共优,毛竹林以毛竹(93.34)占绝对优势,灌木林则主要由经济树种杨梅Myrica rubra(38.99)组成。
3.2 公益林群落的结构
3.2.1 径级结构 中部地区各公益林群落乔木密度大小不一,因此采用百分比表示各径级水平上的分布情况更为直观,中部地区乔木的平均胸径为10.2 cm。由图3A可知:乔木树种主要分布于5 cm≤胸径<10 cm径级,平均为60.53%,径级分布于10 cm≤胸径<15 cm的平均为26.39%,径级分布于15 cm≤胸径<20 cm的平均为8.62%;径级分布于20 cm≤胸径<25 cm的平均为3.01%;径级分布于胸径≥25 cm的平均为1.45%。在各群落中径级为5 cm≤胸径<10 cm的以阔叶林的比例最高,达到73.96%,径级分布于10 cm≤胸径<15 cm的以阔叶林最低16.48%。因此,群落平均胸径表现为杉木林(11.2 cm)>松林(10.6 cm)>针阔混交林(10.2 cm)>毛竹林(9.3 cm)>阔叶林(9.0 cm)>灌木林(8.9 cm)。各公益林群落径级结构虽有不同,却表现一致的趋势,均呈现逆 “J”型,即随着胸径的增大,乔木树种数量的比例逐渐减小。这是由于人类对中部地区森林资源的过度利用,原始的森林植被遭到巨大的破坏,仅留下少量大径级的树木。
3.2.2 垂直结构 中部地区公益林的平均高度H为7.3 m,其高度在6 m≤H<10 m的占主要部分,平均为58.58%,其次为高度在2 m≤H<6 m的占30.34%,高度在10 m≤H<14 m的占18.28%,高度在14 m≤H<18 m的占2.67%,高度在10 m≤H<14 m的占0.13%。各公益林群落中,高度在2 m≤H<6 m的以灌木林最高,占81.92%,高度在2 m≤H<6 m的以松林最高,占53.38%,高度在6 m≤H<10
m和10 m≤H<14 m的均以毛竹林最高,分别占60.02%和8.44%(图3B)。因此,群落平均树高表现为毛竹林(10.6 m)>杉木林(7.5 m)=松林(7.5 m)>针阔混交林(7.1 m)>阔叶林(6.4 m)>灌木林(4.3 m),松林、杉木林、针阔混交林、阔叶林等4种公益林群落垂直结构差异不大,毛竹林高度分布呈纺锤型,其高度分布较为合理。

表3 浙江中部地区公益林主要植物种重要值Table 3 Important values of main plant species in the central Zhejiang

图3 浙江中部地区公益林径级(A),垂直结构(B)和林龄结构(C)Figure 3 Structure of diameter(A),vertical(B)and age(C)in the central Zhejiang
3.2.3 林龄结构 群落的年龄结构决定着群落的径级和垂直结构。本研究公益林的林龄A为小班内优势树种的平均年龄,取3棵优势树种的平均木,用生长锥测量,辅以公益林二类资源清查数据对照,结果较为一致。由图3C可知:毛竹林的林龄均小于10年生,这是由于毛竹林在经营上多为隔年留养新竹,隔年采伐老竹。一般6年生老竹就采伐,因而现存毛竹林常是1,3,5年生类型或2,4,6年生类型。除毛竹林外,林龄(A)在A<10 a和10 a≤A<20 a比例以灌木林最大,分别为47.62%和35.71%。林龄在20 a≤A<30 a之间的以针阔混交林最大,分别为 49.43%和林龄在30≤A<40的以松林最大,为29.27%,在40 a≤A<50 a的以阔叶林最大,为13.24%。因此平均林龄依次为阔叶林(26.1 a)=针阔混交林(26.1 a)>松林(24.0 a)>杉木林(23.3 a)>灌木林(12.1 a)>毛竹林(4.4 a),松林、 杉木林、 针阔混交林、阔叶林4种公益林群落年龄结构呈现纺锤型分布,且差异不大,表明样方的选择在林龄上较为一致。
3.3 优势种群静态生命表
乔木层的优势种往往是群落的建群种,其种群结构直接影响着群落的结构。为此,本研究对中部地区各公益林群落主要有优势种马尾松、杉木(演替初期先锋种)、木荷(演替中期优势种)、青冈(演替后期优势种)[22]编制静态生命表(表4),4 个种群密度分别为 476 株·hm-2,260 株·hm-2,102 株·hm-2和 34株·hm-2(毛竹径生长在1 a内完成,杨梅为经济树种,故在此不作分析)。从各龄级的存活数和存活率上看,优势种群随龄级的增大,其存活数和存活率均依次减少,种群在龄级Ⅰ和Ⅱ个体数量丰富,龄级Ⅲ个体数较多,而龄级Ⅳ和Ⅴ个体数量下降迅速,为典型的增长型结构;从生命期望来看,在龄级Ⅰ~Ⅴ范围内,马尾松、杉木作为演替初期的先锋种群其生命期逐渐下降,其中木荷作为演替中期优势种群其生命期望先增后减;青冈的生命期望持续上升,这表明青冈种群作为演替后期优势种更具有保持种群增长的趋势。浙江中部的杉木林多由人工营建而来,少数经砍伐后萌条更新而成。对于杉木各龄级的生命期望,在龄级Ⅲ和Ⅳ略有上升,为波动式的下降,这与杉木个体寿命较长,同时也与杉木萌条的繁殖方式有关,利于保持杉木种群的稳定性。
对中部地区公益林群落主要优势种群各龄级种群的死亡率与环境阻力之间的关系进行分析,结果表明:两者相关性显著,P值分别为马尾松0.829,杉木0.867,木荷0.765,青冈0.950。曲线拟合结果如图4。种群死亡率随着环境阻力的增大而升高,环境阻力是导致个体死亡的主要原因,在龄级Ⅰ~Ⅳ范围内,马尾松种群随着龄级的增加,其所遇到环境阻力持续增加,导致死亡率逐渐增大,个体数量也相应减少;杉木种群则随着龄级的增加,环境阻力先增后降,在龄级Ⅱ达到最大(Kx=1.32),在龄级Ⅰ为最低(Kx=0.51),死亡率也先增后减;木荷种群也表现出死亡率随环境阻力先增后降的规律,环境阻力和死亡率在龄级Ⅱ达到最大(Kx=1.36,Dx=0.75),在龄级Ⅳ降到最低(Kx=0.82,Dx=0.22);青冈种群则随着龄级的增加死亡率随环境阻力持续减小。也就是说,马尾松和杉木作为先锋树种在演替初期在群落中占有优势,在后期种群逐渐衰退,导致群落结构与环境的改变,为演替中期优势种木荷和后期优势种青冈种群的增长创造了条件,这符合正向演替的趋势。

表4 浙江中部地区公益林主要优势种静态生命表Table 4 Static life of dominant species in central Zhejiang

图4 不同龄级优势种死亡率与环境阻力的关系Figure 4 Relationship between mortality and environmental resistance of dominant species in different age class
4 讨论
中部地区是浙江省的生态建设的重点区域,其公益林质量高于全省整体水平,但于人类对中部地区森林资源的过度利用,原始的森林植被遭到巨大的破坏,仅留下少量大径级的树木,且仍以松林、杉木林为主,2种群落占52.87%,因此,目前浙江中部地区的公益林在组成上具有优势种相对单一,乔木层建群树种少、树种构成相对集中,下木层树种丰富且构成相对合理等特点,物种保育能力和群落稳定性都亟待提升。
通过对主要优势种群静态生命表的分析发现,建群种在群落的构建过程中具有控制作用,不同演替阶段优势种群在生活史对策上存在差异,在龄级增加时对环境阻力诸如光照、水分、种间关系等因子以及这些因子随群落正向演替而变化时表现出不同耐受能力。同时,中部地区频繁的人为经营,使得人工林、天然林、天然次生林混杂交错,尤其是人工林往往经历了经营(择伐、皆伐、抚育等)—封育的过程。本研究中,马尾松个体在松林、杉木林、针阔混交林、阔叶林、毛竹林、灌木林中的分布比例为48.19%,3.23%,46.59%,1.11%,0.40%,0.47%;而杉木在各群落中的分布比例为5.90%,41.69%,48.12%,2.14%,1.53%,0.63%。表明在人为和自然因子的干扰下,马尾松和杉木2个种群均与其他树种存在较高程度的混交。因此在龄级Ⅳ时,各种群的环境阻力为马尾松>杉木>木荷>青冈。而这一过程的进展,加速了群落结构的变化,促进了群落演替的发生,可见群落结构的特征是优势种群生活史对策和群落结构变化相互作用的结果。但由于阔叶林的建群种木荷、青冈等种群目前密度很低,且在龄级较低阶段生命期望相比演替先锋种群不占优势,在径级和高度上与其他群落存在差距。因此在今后公益林的经营中注重对优势种群的抚育,对改造和提升阔叶林的质量,加速群落正向演替尤为重要。
[1]宋永昌.生态学·植被生态学[M].上海:华东师范大学出版社,2001.
[2]樊后保.格氏栲群落的结构特征[J].林业科学,2000,36(2):6-12.FAN Houbao.Structure features of Castanopsis kawakamii community[J].Sci Silv Sin,2000,36(2)∶6-12.
[3]达良俊,杨永川,宋永昌.浙江天童国家森林公园常绿阔叶林主要组成种的种群结构及更新类型[J].植物生态学报,2004,28(3):376-384.DA Liangjun,YANG Yongchuan,SONG Yongchuang.Population structure and regeneration types of dominant species in an evergreen broadleaved forest in Tiantong national forest park,Zhejiang Province,Eastern China[J].Acta Phytoecol Sin,2004,28(3)∶376-384.
[4]李亮,刘海丰,白帆,等.东灵山 4种落叶阔叶次生林的物种组成与群落结构[J].生物多样性,2011,19(2):243-251.LI Liang,LIU Haifeng,BAI Fan,et al.Species composition and community structure of four deciduous broadleaved secondary forest in Dongling Mountain[J].Biodiversity Sci,2011,19(2)∶243-251.
[5]JOHNSON J B.Stand structure and vegetation dynamics of a subalpine wooded fen in Rocky Mountain National Park,Colorado[J].J Vegetation Sci,1997,8(3)∶337-342.
[6]HARPER J L.Population Biology of Plants[M].New York∶Academic Press,1977:513.
[7]王鹏,陈丽华,卞西陈,等.北沟林场天然次生林群落结构分布格局[J].应用生态学报,2011,22(7):1668-1674.WANG Peng,CHEN Lihua,BIAN Xichen,et al.Community structure and distribution pattern of a natural secondary forest in Beigou Forest Farm[J].Chin J Appl Eool,2011,22(7):1668-1674.
[8]ARIST M.The structure and dynamics of an Abies pinsapo forest in southern Spain[J].For Ecol Manage,1995,74(1/3):81-89.
[9]AGREN J,ZACKRISSON O.Age and size structure of Pinus sylvestris populations on mires in central and northern Sweden[J].J Ecol,1990,78(4):1049-1062.
[10]CORNELISSEN J,WERGER M,ZHANGCHENG Z.Effects of canopy gaps on the growth of tree seedlings from subtropical broad-leaved evergreen forests of southern China[J].Plant Ecol,1994,110(1):43-54.
[11]杨永川,达良俊.丘陵地区地形梯度上植被格局的分异研究综述[J].植物生态学报,2006,30(3):504-513.YANG Yongchuan,DA Liangjun.A brief review of studies on differentiation of vegetation pattern along a topographic gradient in hilly regions[J].J Plant Ecol,2006,30(3):504-513.
[12]DU Yanjun,MI Xiangcheng,LIU Xaojuan,et al.The effects of ice storm on seed rain and seed limitation in an evergreen broad-leaved forest in east China[J].Acta Oecolog,2012,39∶87-93.
[13]余树全.浙江淳安天然次生林演替的定量研究[J].林业科学,2003,39(1):17-22.YU Shuqian.The quantitative study on the secondary forest succession in Chun’an County,Zhejiang Province[J].Sci Silv Sin,2003,39(1)∶17-22.
[14]袁位高.浙江省生态公益林主要群落结构的比较研究[D].北京:中国林业科学研究院,2009.YUAN Weigao.Comparative Studies on Structure of Main Forest Type of Ecological Service Forest in Zhejiang Province[D].Beijing∶Chinese Forestry Academy,2009.
[15]江波.浙江省生态公益林群落结构特征及其调控研究[D].北京:北京林业大学,2005.JIANG Bo.Studies on Character of Community Structure of the Ecological Forest and Its Quality Control in Zhejiang Province[D].Beijing∶Beijing Forestry University,2005.
[16]张骏.中国中亚热带东部森林生态系统生产力和碳储量研究[D].杭州:浙江大学,2008.ZHANG Jun.NPP and Carbon Storage in Subtropical Forest,Eastern China[D].Hangzhou∶Zhejiang University,2008.
[17]赵常明,陈庆恒,乔永康,等.青藏高原东缘岷江冷杉天然群落的种群结构和空间分布格局[J].植物生态学报,2004,28(3):341-350.ZHAO Changming,CHEN Qinghuan,QIAO Yongkang,et al.Structure and spatial pattern of a natural Abies faxoniana population on the easten edge of Qinghai-Tibetan Plateau[J].Acta Phytoecol Sin,2004,28(3)∶341-350.
[18]熊能,金则新,顾婧婧,等.千岛湖次生林优势种种群结构与分布格局[J].生态学杂志,2010,29(5):847-854.XIONG Neng,JIN Zexin,GU Jingjing,et al.Population structure and distribution pattern of dominant tree species in a secondary forest on Laoshan Island of Qiandao Lake region[J].Chin J Ecol,2010,29(5)∶847-854.
[19]熊能,金则新,陈琢.千岛湖姥山马尾松种群结构和分布格局研究[J].植物研究,2010,30(5)∶537-542.XIONG Neng,JIN Zexin,CHEN Zhuo.Population structure and distribution pattern of Pinus massoniana on Laoshan Island of Qiandao Lake region[J].Bull Bot Res,2010,30(5)∶537-542.
[20]胡小兵,于明坚,陈玉宝.浙北青冈林群落结构与青冈种群数量特征[J].植物研究,2002,22(4):432-438.HU Xiaobing,YU Mingjian,CHEN Yubao.The community structure and the quantity characteristics of Cyclobalanopsis glauca in the northern regions of Zhejiang Province[J].Bull Bot Res,2002,22(4)∶432-438.
[21]王电杰,温国胜,童再康,等.浙江省大鹿岛木麻黄种群静态生命表分析[J].内蒙古农业大学学报:自然科学版,2011,32(4):145-150.WANG Dianjie,WEN Guosheng,TONG Zaikang et al.Analysis to the static life table of Casuarina equisetifolia populations on the big deer island in Zhejiang[J].J Inn Mongolia Agric Univ Nat Sci Ed,2011,32(4)∶145-150.
[22]余树全.浙江省常绿阔叶林的生态学研究[D].北京:北京林业大学,2003.YU Shuquan.The Ecologi cal Studies on Evergreen Broad-leaved Forest in Zhejiang Province[D].Beijing:Beijing Forestry University,2003.
