百山祖中山中亚热带常绿阔叶林群落5年动态特征
2013-05-30陈小荣陈圆圆骆争荣丁炳扬
陈小荣,陈圆圆,骆争荣,丁炳扬
(1.浙江凤阳山-百山祖国家级自然保护区 百山祖管理处,浙江 庆元 323800;2.温州大学 生命与环境科学学院,浙江 温州 325035;3.丽水学院 生态学院,浙江 丽水 323000)
森林并不是一直处于静止不变的环境之中,在大的时间尺度上,伴随着环境气候的变化森林组成的改变是非常明显的[1-2]。森林内主要树种的更新动态,是群落内各种生物和非生物因素综合作用的外在体现,同时对于群落的结构和功能稳定或变化起着十分重要的作用[3-4]。因此,揭示主要树种的更新机制对于预测森林未来的变化趋势,以及深入分析森林更新动态和物种多样性的维持机制具有重要的参考意义。然而,对于成熟的森林来说,其结构和功能都处在一个相对稳定的动态平衡之中,要在短时间内研究成熟森林的动态变化是比较困难的。目前,中国学者对森林更新动态的研究主要集中在种子库和幼苗库动态[4-8]、萌生更新[9]、 林隙下更新[10-11]以及主要群落主要树种的径级结构[5,12-14]等方面。 这些研究往往都是以静态数据(1次调查)来预测森林的动态过程,对于森林动态的研究大多通过间接证据。由于森林动态周期较长,近年来越来越多的学者开始强调自然森林群落长期重复研究的重要性[15-20]。森林动态样地的建立,为分析种群和群落更新动态、物种共存机制、群落水平的演替格局等提供了极有价值的信息[15,21]。目前,国内外已有研究者通过分析动态样地的多次监测数据,研究群落的更新特征[22-23]。百山祖自然保护区主峰北坡1300~1700 m的中山地带保存的中亚热带天然植被长久未受较明显的人为干扰,是研究中山地带亚热带常绿阔叶林群落动态的十分理想的材料[24]。本研究通过分析在百山祖建立的5 hm2的动态监测样地(以下简称“百山祖样地”)2003年和2008年2次全面调查期间群落整体及其中主要物种的短期补员、死亡、适合度及种群大小和群落物种组成的变化情况,了解该地区常绿阔叶林植物种群和群落动态。
1 研究地自然概况
研究地位于浙江省凤阳山-百山祖国家级自然保护区百山祖主峰北坡(27°40′54″N,119°03′53″E)。所在区域属于中亚热带季风气候,受夏季风影响较大,具有明显的季节性变化,年平均气温为12.8℃,年降水量2341.8mm,相对湿度84%,极端最高气温为32.0℃,极端最低气温为-13.0℃。保护区内地形复杂,山地坡度大多在30°以上,海拔800 m以下为红壤,800 m以上为黄壤,棕黄壤集中分布在海拔1700 m的百山祖南坡,地貌类型为深切割中山,以侵蚀地貌为主。
中亚热带常绿阔叶林是百山祖自然保护区的地带性植被。研究样地位于主峰北坡海拔约1400~1600 m的中山地带,由于人为干扰相对较少,该地段至今保留着一些成熟的中国东部亚热带中山典型的常绿阔叶林。样地内植物种类丰富,自然分布的种子植物有77科157属,木本植物和草本植物共305种,其中木本植物181种中包括常绿阔叶物种90种,落叶阔叶物种64种,针叶物种4种,藤本或藤状木本植物23种。该样地中,中山特征性物种短尾柯Lithocarpus brevicaudatus,多脉青冈Cyclobalanopsis multinervis,尖连蕊茶Camellia cuspidata分别替代了该地区低海拔常绿阔叶林常见的石栎Lithocarpus glaber,青冈Cyclobalanopsis glauca,毛花连蕊茶Camellia fraterna等,而苦槠Castanopsis sclero-phylla和红楠Machilus thunbergii没有出现。
2 研究方法
2.1 样地设置
百山祖样地始建于2002年。样地形状为250 m×200 m (5 hm2)的矩形,整个样地建设的方法参照美国Smithsonian热带森林研究中心的建设方案进行[24]。
2003年进行首次群落调查时,对5 hm2样地内所有的乔木和高度2.0 m以上或的胸径≥1 cm灌木编号,进行每木调查,测定内容包括:植物种名、胸径、坐标位置、树高、冠幅和生长状况,并对其标记挂牌,进行长期监测。2008年夏对样地进行复查,内容包括所有2003年调查过的活立木之胸径和生长状况,对新增的胸径到达1.0 cm而未挂牌的树木则还要编号挂牌,并调查植物种名、坐标位置、树高和冠幅。
2.2 数据处理和统计方法
为统计和比较方便,本研究仅分析由2次调查中胸径大于1.0 cm的个体组成。以重要值排在前30位的树种[24]作为研究种群死亡率和补员率主要研究对象。用标准的方法计算死亡率、补员率、种群数量的变化[23]以及相对适合度[25]。通过计算各个物种在不同径级的死亡率(M)和补员率(R)参数,比较物种间的差异。M=(lnN0-lnSt)/T,R=(lnNt-lnSt)/T,λ=R-M,f=R/M 。 其中:T为2次调查时间间隔,N0和Nt分别为某一种群在第1次和第2次调查时的生活个体数,St是该物种在第1次和第2次调查时的都存在生活个体数;λ是种群大小变化率,采用补员率和死亡率的差值来表示;f为物种相对适合度,用补员率和死亡率的比值来表示。死亡个体是指在复查时找到后发现已经死亡或无法找到的植株;补员个体是指在第2次调查中新出现的胸径≥1.0 cm的个体;补员率被定义为数量增长速率常数。
计算某一径级的死亡率和补员率时,当N0=0且St=0时,死亡率M=0;当Nt=0且St=0时,补员率R=0;当N0≠ 0且St=0时,死亡率M=lnN0/T;当Nt≠ 0且St=0时,补员率R=lnNt/T。同时,根据样地内木本植物径级结构特点,采用上限排除法将不同树种归入乔冠层、乔亚层、小乔木和灌木共4个功能型层次。各功能型的径级划分标准为乔冠层和乔亚层:Ⅰ级(1 cm≤胸径<2.5 cm),Ⅱ级(2.5 cm≤胸径<7.5 cm),Ⅲ级(7.5 cm≤胸径<22.5 cm),Ⅳ级(胸径≥22.5 cm);小乔木:Ⅰ级(1.0 cm≤胸径<2.5 cm),Ⅱ级(2.5 cm≤胸径<5 cm),Ⅲ级(胸径≥5 cm);(3)灌木:Ⅰ级(1.0 cm≤胸径<2.0 cm), Ⅱ级(2.0 cm≤胸径<3.0 cm), Ⅲ级(胸径≥3.0 cm)。
为了研究群落在2003年和2008年物种组成的变化,本研究将整个5 hm2样地平均分割为4个1.25 hm2的区块(125 m×100 m,即左下角区块、右下角区块、左上角区块和右上角区块),以群落物种组成的空间差异作为参照,评价群落物种组成在时间上的变化程度。采用非计量多元尺度法(NMS)排序提取群落物种组成差异,排序轴的显著性评估用Monte carlo检验。本分析通过R软件平台中的Smacof软件包实现。
3 结果
3.1 死亡和补员个体的分布
从2003年到2008年,该样地内胸径≥1.0 cm的植物个数从30092株下降到28880株,共有2343株死亡,占样地内个体总数的7.79%,分属于96个物种,占物种总数62.75%。其中,乔木57种,灌木22种,藤本17种。死木中胸径最大为40.0 cm(短尾柯),最小为1.0 cm。
样地内死亡个体数最多的物种是麂角杜鹃Rhododendron latoucheae,为417株,其次为窄基红褐柃Eurya rubiginosa var.attenuote和尖连蕊茶,分别为357和305株。其余物种个体死亡数在100株以上的还有水丝梨Sycopsis sinensis、短尾柯、厚叶红淡比Cleyera pachyphylla。此外,已有7个物种从样地中消失, 皆为稀有种(≤1株·hm-2)。
5 a间,新增加的胸径≥1.0 cm的木本植物共有1131株,形成补员群体,占样地内个体总数的3.93%。有补员个体的物种共计65种,占物种总数的36.72%,其中乔木46种,灌木15种,藤本3种。
样地内麂角杜鹃和尖连蕊茶的补员个体数最多分别为185株和154株,其次为窄基红褐柃、水丝梨、厚叶红淡比,分别为108,107株和103株。补员个体数在100株以上的物种有5种,除了窄基红褐柃以外,其余4种的重要值在所有物种中居前10位。
3.2 主要树种的死亡率和补员率
将30种主要树种按照生活型分为22种乔木和8种小乔木或灌木。样地内30个主要树种胸径≥1.0 cm的个体5年内的年平均死亡率为1.55%,年平均补员率为0.78%。从整个种群来比较,30个树种的整体死亡率、补员率以及适合度差异较大,死亡率和补员率跟物种多度之间并没有显著的相关性(表1),多度高的物种也没有显示出更高的相对适合度。

表1 百山祖常绿阔叶林30个主要树种的补员率、死亡率和适合度Table1 Mortality,recruitment rates and fitness of 30 important tree species in Baishanzu evergreen broad-leaved forest
30个主要物种的死亡率比较结果显示:中华石楠Photinia beauverdiana的死亡率最高,年平均达3.49%;亮叶水青冈Fagus lucida的死亡率最低,年平均仅为0.79%。死亡率排名前10位的物种中,只有窄基红褐柃和水丝梨的个体密度较大;而中华石楠,合轴荚蒾Viburnum sympodiale,厚皮香Ternstroemia gymnanthera,交让木Daphniphyllum macropdum的相对多度值并不大,但死亡率却很高。此外,麂角杜鹃,多脉青冈,木荷Schima superba的重要值分别位列第1位、第7位和第10位,但死亡率却远远低于平均值,这些物种在5 a间均保持着优势。
30个主要物种的补员率比较结果显示:四川山矾Symplocos lucida的补员率最大,年平均高达3.35%,其次为浙闽新木姜子Neolitsea aurata var.undulatula,水丝梨,凸脉冬青Ilex editicostata,台湾冬青Ilex formosana年平均补员率分别为1.55%,1.35%,1.33%和1.32%。相比之下,石灰花楸Sorbus folgneri,合轴荚蒾,云锦杜鹃Rhododendron fortunei,微毛柃Eurya hebeclados等的补员率很低,年平均都不到0.50%。
3.3 死亡率和补员率与功能型的关系
百山祖5 hm2样地,胸径≥1.0 cm的所有个体的年死亡率为1.45%、年补员率为0.62%。在按照生活型划分出4个功能群的死亡率中,灌木的年平均死亡率超过2.0%,高于其他3个功能群(图1)。4个功能群中,乔亚层平均补员率最高超过了1.0%,而小乔木的平均补员率最低,乔冠层、灌木层位居中间,但总体来说,4个功能群之间的平均补员率差距相对较小。
由表2可见,除乔冠层以外,补员率均随着径级的增大而减少;乔冠层和小乔木径级Ⅰ补员数明显多于死亡数,其余径级的补员数均少于死亡数。乔冠层物种和乔亚层物种在径级I平均死亡率最高,在径级Ⅱ和径级Ⅲ平均死亡率随着径级的增大而减少,而在径级Ⅳ略有上升,在不同径级间的变化差异比较大;小乔木和灌木表现为平均死亡率随着径级的增大而减少。

表2 4个功能群死亡率和补员率的径级分布Table2 Mortality and recruitment rate of four different growth forms in each DBH class
3.4 种群大小变化趋势
物种补员与死亡数目的差值,反映出该种群大小的变化,而补员率与死亡率的差值,则表征种群的变化率。30个主要树种中,只有四川山矾1个物种的补员数目超过死亡数目,种群大小的变化率为1.25%,表明该种群规模在扩大。有28个物种的种群大小在下降,但多数种群降幅并不大,种群变化率为-0.04%~-3.16%,其中合轴荚蒾(-3.16%)和树参Dendropanax dentiger(-2.52%)的降幅最大。黄山木兰Magnolia cylindrica的种群大小没有发生变化。
3.5 群落物种组成的变化
NMS排序发现样地的4个区块明显存在物种组成差异,NMS提取2条排序轴,压力系数(stress)=0.0015,P<0.01。排序结果显示:4个区块的分布在排序图的四边,并且区块空间上距离越远,在排序图上的距离也越。在2次调查间,4个区块的物种组成发生了不同程度的变化,并且呈现出相似的变化趋势(图2)。从图2可以看到:4个区块2次调查间的差异远小于它们之间由空间造成的物种组成差异。
4 讨论
4.1 群落的总体动态

图1 不同群落层次的死亡率和补员率Figure1 Mortality and recruitment rate of different community-level
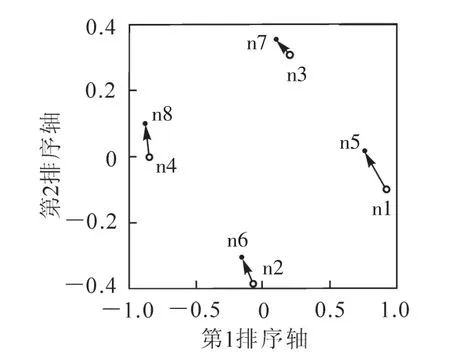
图2 群落不同区块2次全面调查物种组成非计量多元尺度法(NMS)排序Figure2 NMS ordination for tree composition of 4 subplots in 2003 and 2008
本研究结果显示,在百山祖样地,胸径≥1.0 cm的所有个体的年死亡率为1.45%,年补员率为0.62%;30个主要物种的平均年死亡率和平均年补员率为1.55%和0.78%。巴拿马Barro Colorado(BCI)样地所有胸径≥1.0 cm个体的年死亡率为2.64%,年补员率为2.87%,马来西亚Pasoh样地则分别为1.46% 和1.65%[23]。 日本4 hm2暖温带常绿阔叶林中这2个值分别为1.45% 和2.44%[26]。 古田山5 hm2样地则分别为2.02%和5.09%,43个主要物种的平均年死亡率和平均年补员率分别为2.26%和5.90%[22]。可见,百山祖样地的总体死亡率处于相对较低的水平,与日本的暖温带常绿阔叶林接近。
群落的总体死亡率受多种因素影响。一般来说个体死亡率与个体的径级有关,径级较小的个体容易死亡[22,26-28]。本研究结果也表明不同功能群不同径级的死亡和补员率各异。总体上,各功能群植物的平均死亡率随着径级的增大而下降。然而,百山祖样地的森林群落处于群落演替的高级阶段,总体上群落内小个体相对较少,同时,受外界干扰和自然灾害较少。这就使百山祖样地群落整体死亡率较低。另外,与其他地点的研究比较发现,百山祖样地群落的补员率也远低于其他样地。百山祖样地的木本植物个体在2003-2008年间数量呈现下降趋势,降幅达4.20%。随着群落总个体数的下降,已有7个稀有种从样地中消失。本样地的其他研究人员在监测样地部分区域近4 a幼苗的变化时也发现,监测区域的幼苗无论是个体数还是种数、科属数都有减少的趋势。我们于2008年至今对百山祖样地的幼苗动态监测(3次·a-1)也显示,样地内的幼苗也出现了类似的减少趋势。这说明目前的群落生境条件已经使群落的整体更新出现了一定的困难(施璐璐等,待发表)。群落动态变化趋势受到存活、补充和死亡树木个体变化的影响。骆争荣[29]对百山祖样地被监测30个常见树种进行了基于个体的存活分析和基于种群的密度制约稀疏效应空间格局分析,结果表明密度制约效应仅对少数物种存在作用,且随着群体年龄的增长密度制约的稀疏效应的强弱和有无会发生变化,密度制约的稀疏效应对种群的调节不是一贯的。因此,由密度相关自疏效应对于百山祖亚热带常绿阔叶林的结构和动态影响是微弱的。然而,群落自身的随机波动,群落对全球气候变化的响应,或样地建立后引入了一定的人为干扰都会影响群落的更新。到底是何种机制造成了这种现象还有待于进一步的数据积累和研究。
4.2 种群大小的变化
Condit等[23]的研究表明,Barro Colorado岛由于经历过严重干旱所以其样地内10%的物种变化率超过5%,而对比干扰较小的Pasoh样地只有2%的物种变化率超过5%。在古田山的研究发现由于受生境异质性影响,样地内43个重要种群中有10种群的变化率超过5%,比例达23.26%[22]。 在百山祖样地,我们没有发现种群变化率超过5%的物种,只有10%的物种变化率超过2%,分别为中华石楠、树参、合轴荚蒾。同为亚热带常绿阔叶林,百山祖样地内种群的变化程度没有古田山样地迅速,群落相对稳定。同样为5 hm2的面积,古田山样地仅有16812个胸径≥1.0 cm的木本植物个体[22],而百山祖样地胸径≥1.0 cm的木本植物个体数达到了30092个。较高的植株密度使样地内任何物种都很难在短时间内实现种群的较大增长;同时,由于百山祖北坡的常绿阔叶林为保存完好的天然植被[24],群落内部物种竞争关系比较稳定,竞争排斥实际上较难发生,且人为干扰较少,因此种群也不会发生急剧缩减。群落物种组成的稳定性也反映了种群大小的低变化率。非计量多元尺度法(NMS)分析结果表明,从2003年到2008年虽然群落物种组成发生了一些变化,但这些变化相对于群落物种组成的空间差异小得多,群落物种组成相对稳定。
4.3 种群补员率、死亡率和适合度的种间差异
和基于物种生态位分化的传统生态位理论不同,近年来在群落生态学界受到广泛研究的中性理论认为,群落中所有物种的个体都具有完全相同的出生率、死亡率和扩散率,即物种具有生态等同性[25,30]。然而,我们发现在百山祖样地的30个主要树种间的死亡率和补员率存在着较大的差异,并没有显示出如中性理论所说的物种生态等同性。Hubbell[30]声称生态位分化可能使得物种的相对适合度相同。这里我们使用一个个体在其生活史中繁殖的新个体的数量来表示植物的相对适合度。结果显示,百山祖常绿阔叶林30个主要树种的相对适合度在0~1之间变化,显示出了较大的种间差异。因此,百山祖常绿阔叶林中的大多数树种看起来并不是中性的。
百山祖样地群落整体的年死亡率和年补员率都较30个主要物种的平均年死亡率和平均年补员率低。这说明个体数较少的物种(小种群种)相对于主要物种在存活上有一定优势,这可能是小种群种能在本群落中维持的重要原因。但是小种群种也体现出了比主要种更小的补员率,说明小种群种在群落中遇到的更新障碍比主要种更大,种群更新困难。因此,从总体上来说,主要种比小种群种具有更高的种群更替速率。
从死亡和补员的数量来看,补员和死亡个体数目在物种间分布不均匀,随物种的不同而变化;排名前几位的都是该群落的优势物种,如麂角杜鹃、尖连蕊茶、窄基红褐柃、水丝梨和厚叶红淡比等。乔冠层树木、乔亚层树木、小乔木、灌木等4种生活型中灌木的平均死亡率超过了2%,乔亚层的平均补员率则表现为最高。但是各物种的死亡率和补员率和多度并没有显著的相关性,多度高的物种也没有更高的相对适合度。因此,从死亡率和补员率来看,优势种并没有体现出明显的存活优势和更新优势。
在植物一生的生长进程中,种子扩散、萌发和植株生长、存活等过程都会受到各种因素的制约。在该样地开展的幼苗动态监测也显示,在郁闭的林下层,由于阳光不易到达,成为幼苗、幼树更新和存活的最主要限制因素(施璐璐等未发表数据)。光耐受性成为植物更新的重要条件之一[31]。在4种功能型中,乔冠层和乔亚层在小径级时的死亡率最高,小乔木的死亡率最低,其原因可能就在于百山祖样地乔冠层和乔亚层树种多数不是光耐受型,所以在小径级的时期由于光照不足而容易死亡。
[1]BUSH M B, COLINVAUX P A, WIEMANN M C, et al.Late pleistocene temperature depression and vegetation change in Ecuadorian Amazonia[J].Quaternary Res, 1990, 34(3):330-345.
[2]PIPERNO D R.Vegetational history of a site in the central amazon basin derived from phytolith and charcoal records from natural soils[J].Quaternary Res, 1996, 45:202-209.
[3]王利伟,李步杭,叶吉,等.长白山阔叶红松林树木短期死亡动态[J].生物多样性,2011,19(2):260-270.WANG Liwei, LI Buhang, YE Ji, et al.Dynamics of short-term tree mortality in broad-leaved Korean pine (Pinus koraiensis) mixed forest in the Changbai Mountains[J].Biodiversity Sci, 2011, 19(2):260-270.
[4]陈小勇,宋永昌.洪水干扰对青冈种群的影响[J].热带亚热带植物学报,1997,5(1):53-58.CHEN Xiaoyong,SONG Yongchang.Influence of flood disrutbance on the regeneration of Cyclobalanopsis glauca populations[J].J Trop Subtrop Bot, 1997, 5(1):53-58.
[5]谢凡,王素珍.井冈山区常绿阔叶林动态研究[J].应用生态学报,1991,2(1):1-7.XIE Fan,WANG Suzhen.Dynamic studies on regerated seedlings of evergreen broadleaf forest in Jinggangshan re-gion [J].J Appl Ecol, 1991, 2(1):1-7.
[6]彭军,李旭光,付永川,等.重庆四面山常绿阔叶林建群种种子雨、种子库研究[J].应用生态学报,2000,11(1):22-24.PENG Jun, LI Xuguang, FU Yongchuan, et al.Seed rain and seed bank of constructive species in evergreen broadleaved forest at Chongqing Simian Mountain[J].J Appl Ecol, 2000, 11(1):22-24.
[7]田丽.黄土高原马栏林区森林群落更新研究[D].西安:陕西师范大学,2007.TIAN Li.Studies on the Regeneration of Forest Communities in Malan Forest Region on the Loess Plateau [D].Xi’an:Shaanxi Normal University,2007.
[8]张健,李步杭,白雪娇,等.长白山阔叶红松林乔木树种幼苗组成及其年际动态[J].生物多样性,2009,17(4):385-396.ZHANG Jian, LI Buhang, BAI Xuejiao, et al.Composition and interannual dynamics of tree seedlings in broadleaved Korean pine (Pinus koraiensis) mixed forest in Changbai Mountain[J].Biodivesity Sci, 2009, 17(4):385-396.
[9]何永涛,曹敏,唐勇,等.哀牢山中山湿润常绿阔叶林冠层树木萌生现象的初步研究[J].武汉植物学研究,2000,18(1):523-527.HE Yongtao, CAO Min, TANG Yong, et al.A preliminary study on sprouting of canopy trees in middle mountain moist evergreen broad-leaved forest of Ailao Mountain, Yunnan[J].J Wuhan Bot Res, 2000, 18(1):523-527.
[10]臧润国.林隙更新动态研究进展[J].生态学杂志,1998, 17(2):50-58.ZANG Runguo.Research advances of gap regeneration dynamics[J].Chin J Ecol, 1998, 17(2):50-58.
[11]王周平,李旭光,石胜友,等.缙云山针阔混交林林窗期树木更新[J].植物生态学报,2001,25(1):399-404.WANG Zhouping, LI Xuguang, SHI Shengyou, et al.Forming characteristics of gap in the forest in Jinyun Mountain[J].Acta Phytoecol Sin, 2001, 25(1):399-404
[12]于明坚.青冈常绿阔叶林群落动态研究[J].林业科学,1999,35(6):42-51.YU Mingjian.Dynamics of an evergreen broadleaved forest dominated by Cyclobalanopsis glauca in southeast China[J].Sci Silv Sin, 1999, 35(6):42-51.
[13]符婵娟.神农架巴山冷杉群落更新特点及影响因素研究[D].北京:北京林业大学,2010.FU Chanjuan.Regeneration Characteristic and Influencing Factors of Abies fargesii Forests in Shennongjia National Nature Reserve[D].Beijing: Beijing Forestry University, 2010.
[14]陈小荣,李乐,夏家天,等.百山祖亮叶水青冈种群结构和分布格局[J].浙江农林大学学报,2012,29(5):647-654.CHEN Xiaorong, LI Le, XIA Jiatian, et al.Population structure and distribution of Fagus lucida in the Baishanzu forest[J].J Zhejiang A&F Univ, 2012, 29(5): 647-654.
[15]CONDIT R.Research in large, long-term tropical forest plots [J].Trends Ecol Evol, 1995, 10(1): 18-22.
[16]UMEKI K, KIKUZAWA K.Long-term growth dynamics of natural forests in Hokkaido, northern Japan [J].J Veg Sci,1999, 10(3): 815-824.
[17]侯继华,黄建辉,马克平.东灵山辽东栎林主要树种11年种群生长动态[J].植物生态学报,2004,28(1):609-615.HOU Jihua, HUANG Jianhui, MA Keping.Eleven-year population growth dynamics of major species in a Quercus liaotungensis forest in the Dongling Mountains, Northern China[J].Acta Phytoecol Sin, 2004, 28(1):609-615.
[18]LEGENDRE P, MI Xiangcheng, REN Haibao, et al.Partitioning beta diversity in a subtropical broad-leaved forest of China[J].Ecology, 2009, 90: 663-674.
[19]戴景贤.次生阔叶林更新方式对林分生长和演替的影响研究[J].福建林业科技,2007,34(3):60-64.DAI Jingxian.The growth performance and succession of subnatural broadleaved forest affected by regeneration patterns[J].J Fujian For Sci Technol, 2007, 34(3): 60-64.
[20]冯平.人为干扰对米槠群落更新动态的影响[D].福州:福建农林大学,2007.FENG Ping.The Effect of Anthropogenic Interference on the Updating Dynamic of Castanopsis carlesii Community[D].Fuzhou:Fujian Agriculture&Forestry University, 2007.
[21]AUSTIN M P.Permanent quadrats:an interface for theory and practice [J].Plant Ecol, 1981, 46:1-10.
[22]汪殷华,米湘成,陈声文,等.古田山常绿阔叶林主要树种 2002-2007年间更新动态[J].生物多样性,2011, 19(2):178-189.WANG Yinhua, MI Xiangcheng, CHEN Shengwen, et al.Regeneration dynamics of major tree species during 2002-2007 in a subtropical evergreen broad-leaved forest in Gutianshan National Nature Reserve in East China[J].Biodiver Sci, 2011, 19(2):178-189.
[23]CONDIT R, ASHTON P S, MANOKARAN N.Dynamics of the forest communities at Pasoh and Barro Colorado:comparing two 50-ha plots[J].Philo Trans Royal Soc London Series Bio Sci, 2006, 354: 1739-1748.
[24]徐敏,骆争荣,于明坚,等.百山祖北坡中山常绿阔叶林的物种组成和群落结构[J].浙江大学学报:农业与生命科学版, 2007, 33(4): 450-457.XU Min, LUO Zhengrong, YU Mingjian, et al.Floristic composition and community structure of mid-montane evergreen broadleaved forest in north slope of Baishanzu Mountain [J].J Zhejiang Univic Agric&Life Sci, 2007, 33(4): 450-457.
[25]周苏荣,张大勇.群落生态学的中性理论[J].植物生态学报,2006,30(5):868-877.ZHOU Surong, ZHANG Dayong.Neutral theory of community ecology[J].Acta Phytoecol Sin, 2006, 30(5): 868-877.
[26]HOSHINO D, NISHIMURA N, YAMAMOTO S.Dynamics of major conifer and deciduous broad-leaved tree species in an old-growth Chamaecyparis obtusa forest, central Japan[J].For Eco Manage, 2002, 159: 133-144.
[27]KING D A, DAVIES S J, NOOR N S M.Growth and mortality are related to adult tree size in a Malaysian mixed dipterocarp forest[J].For Ecol Manag, 2006, 223:152-158.
[28]LUO Zhengrong, MI Xiangcheng, CHEN Xiaorong, et al.Density dependence is not very prevalent in a heterogeneous subtropical forest[J].Oikos, 2012, 121:1239-1250.
[29]骆争荣.百山祖亚热带常绿阔叶林群落密度制约作用[D].杭州:浙江大学,2011.LUO Zhengrong.The Effects of Density Dependence in Subtropical Evergreen Broad-leaved Forest of Baishanzu[D].Hangzhou:Zhejiang University,2011.
[30]HUBBELL S P.The Unified Neutral Theory of Biodiversity and Biogeography [M].Princeton:Princeton University Press,2001.
[31]梁晓东,叶万辉,蚁伟民.林窗与生物多样性维持[J].生态学杂志,2001,20(5):64-68.LIANG Xiaodong, YE Wanhui, YI Weimin.Gap dynamics and the maintenance of biodiversity [J].Chin J Ecol,2001, 20(5):64-68.
