遮荫干扰对天然羊草分株种群光合产物及其分配的影响
2013-11-05王洪义王智慧
王洪义,王智慧
(黑龙江八一农垦大学,黑龙江 大庆163319)
羊草(Leymus Chinensis (Trin.)Tzvel.)是禾本科(Gramineae)赖草属的一种多年生草本植物,主要分布于欧亚草原带的东部自贝加尔湖到我国的东北平原。它是一种典型的克隆植物,生态适应性好,生态可塑性强,既可以分布在地带性的草甸草原和典型草原,又可群生于非地带性的盐碱地和低洼地段上。与同地带其它植物相比,羊草具有适口性好,非常适合温带草原这种光、温、水的组合效应,而且没有夏枯现象,具有高额的绿色物质产量和优良的饲用质量,是我国主要的出口牧草之一[1-3]。
环境条件的差异是影响植物生长、发育和分布的重要因素。在自然界中,环境差异普遍存在,因而对植物的影响较普遍[4]。光作为重要的环境因子,光照强度的强弱对植物的生长发育有重要影响[5]。遮荫可以模拟光照强度的减弱,研究植物对光强弱变化的反应,目前对于遮荫的研究多见于一些经济作物如蔬菜或者花卉等,且大部分是室内盆栽,如董如磊[6]对野茉莉遮荫后幼苗形态进行研究,刘贤赵[7]对番茄不同生育阶段遮荫对光合作用与产量的影响进行了研究,刘广林[8]对扶芳藤遮荫处理后光合特性进行了研究。而自然条件下从种群或群落水平的遮荫效应研究非常少,尤其是对我国温带草原羊草种群遮荫干扰的研究目前还少见报道。
羊草的光合生产能力和产物的运输分配方式直接影响最终生物产量的形成[9]。也就是说,从光合生产变成较高的生物产量之间,存在一个有机物的运输和分配问题。植物体内有机物的运输主要指光合产物——可溶性糖的形式在植物体内运输[10-11]。因此可以通过可溶性糖的含量变化,了解植物体内的能量分配。羊草分株种群是典型的克隆植物,在环境胁迫下,各分株之间存在一种能量互补及生殖投资权衡关系[12]。本实验选取天然羊草种群进行遮荫研究,目的通过测定光合产物(可溶性糖)的分配变化,研究羊草分株种群在光胁迫干扰下的能量分配以及种群内部的互惠共生和繁殖策略。
1 材料与方法
1.1 材料及研究区概况
试验地点选在中国科学院植物研究所多伦恢复生态学试验示范研究站(内蒙古多伦县境内:E 116o44',N 42o29',Alt.1 458 m)的实验样地,是分布相对均匀的天然羊草单优群落。土壤是沙质壤土,年降雨量是250 ~400 mm,年蒸发量1 600 ~1 800 mm,年均气温1.6 ℃左右,最暖月(7 月)平均气温为18.7 ℃,最冷月(1 月)平均气温 -18.3 ℃,无霜期100 d 左右,≥10 ℃积温1 917.9 ℃。
1.2 试验设计
本实验在5 月初羊草营养生长开始时期进行,选择地势较平坦排水良好的地块,设置4 种遮光梯度处理,遮光强度分别为0%(CK)、30%(S1)、50%(S2)、70%(S3)4种处理,S1 处理采用一层纱网布,S2 和S3处理分别用相当强度的遮阳网。每个小区面积4 m×4 m(图1),为防止各小区间相互影响,小区间有2 m 的通道。每个处理重复5 次,一共20 个试验小区,各小区随机排列。

图1 样地现场布置图Fig.1 Treatment of sample plot
1.3 测定方法
到8 月底种子成熟后,在每个小区中央取0.5 m×0.5 m 的小样方,用剪刀剪掉地上部分,用信封装好,带回实验室后将植株分成穗、茎、叶片、叶鞘、立枯几部分,并按照小区编号进行分装;小样方的地下部分用铁铲沿小样方周围线切下,深度为30 cm,把整个土块用塑料袋装好,回去放在大盆里用水慢慢泡开,用孔径0.5 mm 的纱网把根捞出,再用清水冲洗数次(冲洗时注意须根别流失,根茎别弄断),然后人工将根茎和细根分开,按照小样方的编号将各个部分分装,所有的植物样品均在75 ℃下烘干24 h,直到样品达到恒质量。在实验室中,测定了以下部分可溶性糖测含量:细根、根茎、地上茎(茎)、地下茎、穗、叶片、叶鞘、立枯。
2 结果与分析
在光合产物分配的研究中,根据植株各部分功能不同,将植株分成8 个部分,表1 为各部分在不同处理下的可溶性糖含量变化。从表1 可以看出,在P <0.05 水平上,只有茎、叶鞘、叶片和根茎的含糖量在4 种处理间存在显著差异,其它组分上没有显著差异。各部分含糖量由多到少依次是:地下茎、地上茎、根茎、叶鞘、叶片、穗、立枯和细根,且各处理间糖的分配有一定差异性。
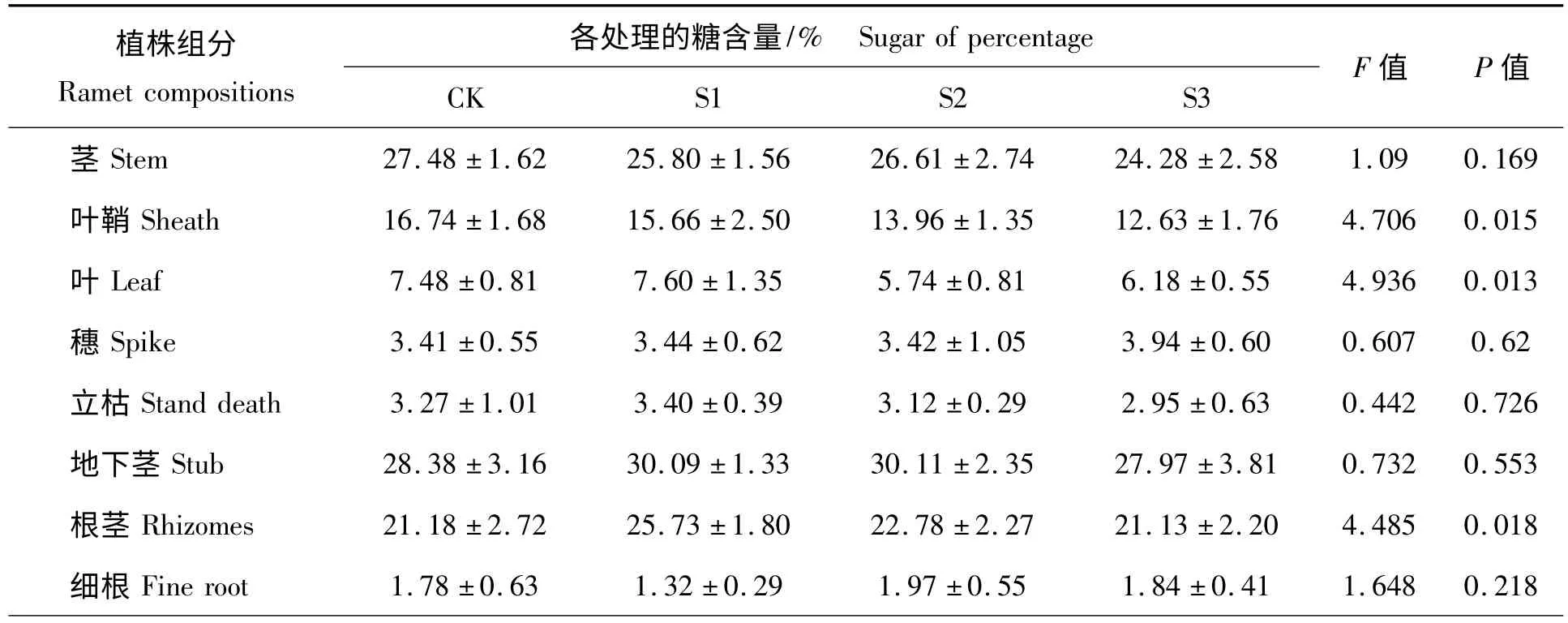
表1 不同处理下植株各部分的含糖量分析Tab.1 Analysis on correlative relationship about sugar of composition of ramet under different shading treatments
不同遮荫处理下,各器官平均生物量变化情况见表2。从表2 分析可以得出,细根生物量最大占整个植株生物量一半以上,但细根的含糖量较低,说明并不是羊草的重要储存器官;S3 处理穗的生物量最大,同时含糖量也较高,说明该处理有利于穗器官的发育;S1 处理地上生物量大于其他处理,其它器官生物量与遮荫之间关系不明显。
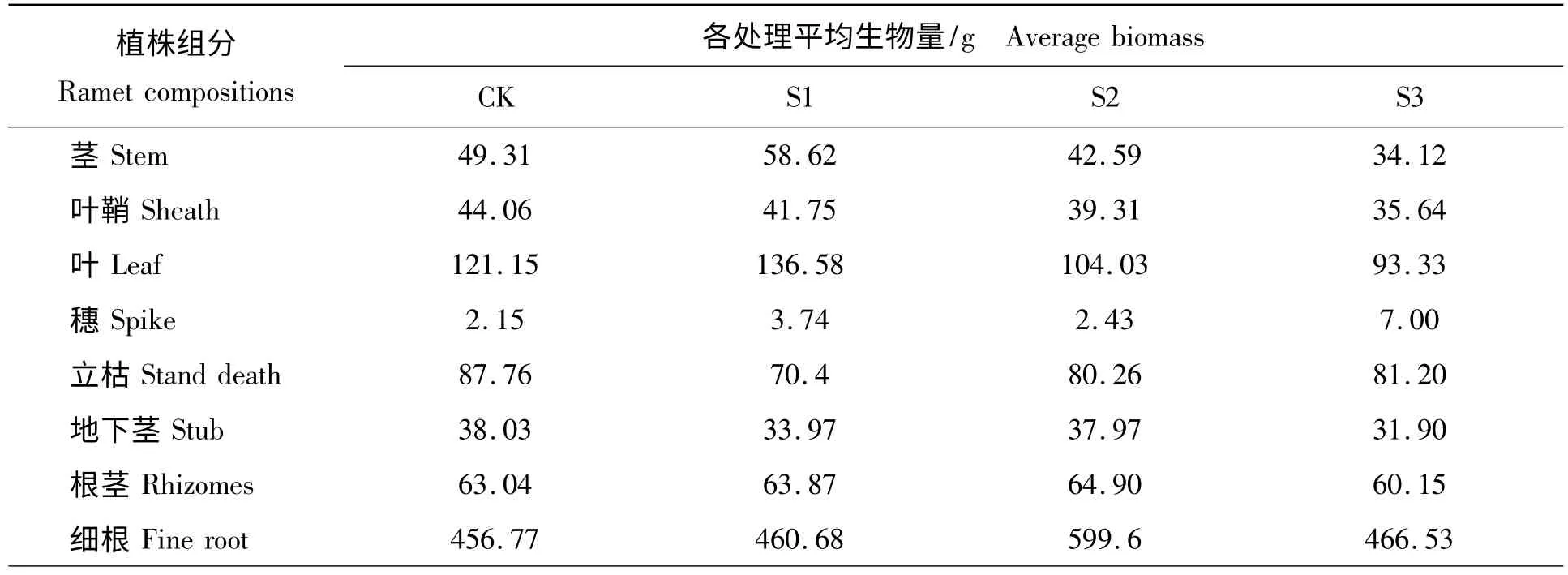
表2 不同处理下植株各部分生物量分析Tab.2 Analysis on correlative relationship about biomass of ramet under different shading treatments
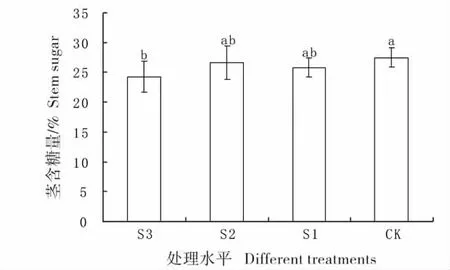
图2 不同遮荫处理下羊草茎的含糖量Fig.2 Percentage content of sugar of stem under different shading treatments
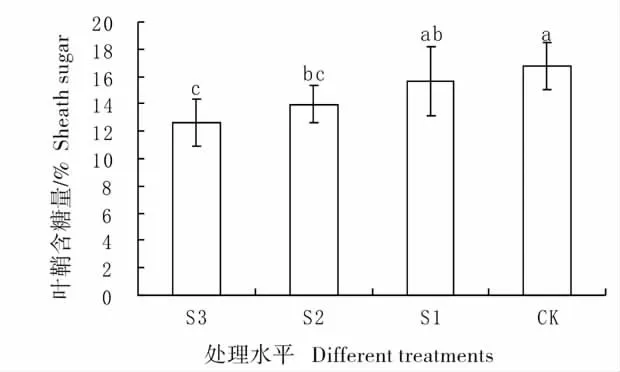
图3 不同遮荫处理下羊草叶鞘的含糖量Fig.3 Percentage content of sugar of sheath under different shading treatments
2.1 不同遮荫处理下羊草地上茎的含糖量比较
图2 为不同遮荫处理下地上茎的含糖量比较,在4 种处理中,CK 组地上茎的含糖量最高,达到25%以上,并且与S3(70%)遮荫有显著差异,表明重度遮荫情况下,羊草地上茎的糖分积累减少;S2 和S1 处理与CK 处理无显著差异。
2.2 不同遮荫处理下羊草叶鞘含糖量的比较
在羊草植株内叶鞘的含糖量也是较高的。羊草叶鞘的含糖量随着遮荫梯度的增加而逐渐降低(图3),CK 组糖含量超过16%,而在S3 处理下,糖含量只有13%,在4 种处理下,CK 与S2、S3 达到显著或极显著差异,S1 与S3 也达到显著差异水平(表1)。说明遮荫可以影响叶鞘器官的含糖量,使糖分出现一定的重新分配。
2.3 不同遮荫处理下羊草叶片含糖量的变化
含糖量最高的是S1 处理(图4),大约7%左右,最低的是S2 处理,大约是5.5%左右,并且S1 和CK 与遮荫较高的S2 和S3 均呈显著差异,说明遮荫情况下,羊草的叶片储存碳水化合物的能力减弱,这可能与遮荫情况下羊草的叶片厚度减少有关[13],减弱了碳水化合物的储存量。轻度遮荫和对照之间无显著差异;中重度遮荫(S2、S3)间也没有显著差异。
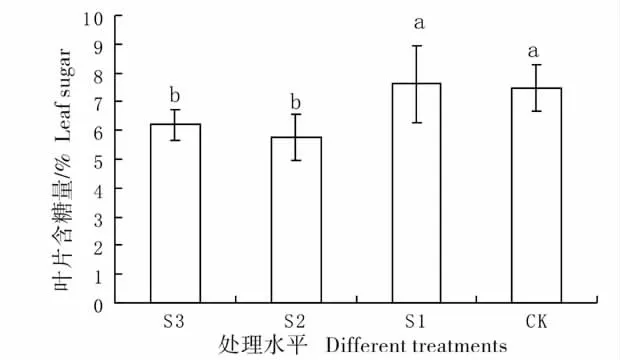
图4 不同遮荫处理下羊草叶片的含糖量Fig.4 Percentage content of sugar of leaf under different shading treatments
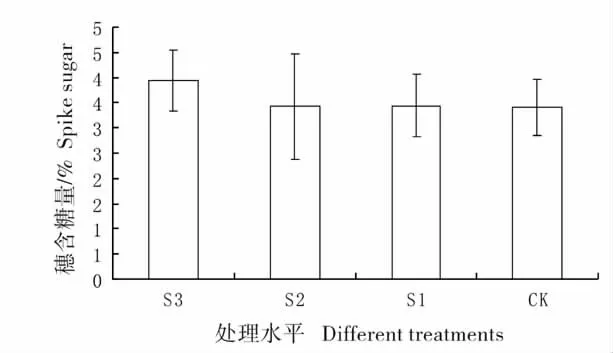
图5 不同遮荫处理下羊草穗的含糖量Fig.5 Percentage content of sugar of spike under
2.4 不同遮荫处理下羊草穗的含糖量变化
S3 的穗含糖量最高(图5),其它三组含糖量基本一样,各组之间并没有达到显著差异,说明遮荫处理并不能显著的改变穗的含糖量。但从图中可以看出,重度遮荫(S3)处理的含糖量增加明显,可以推断此种情况下,羊草进行了有性繁殖投资倾向。
2.5 不同遮荫处理下羊草立枯的含糖量变化
羊草立枯间含糖量并无显著差异(图6),含糖量最高的是S1 处理,含糖量3.5%左右,其它几组含糖量基本相同。
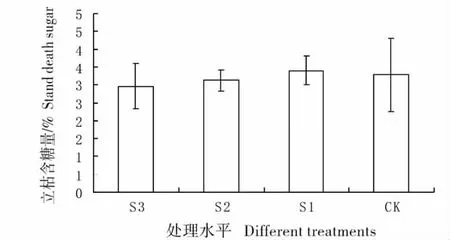
图6 不同遮荫处理下羊草立枯的含糖量Fig.6 Percentage content of sugar of stand death under different shading treatments
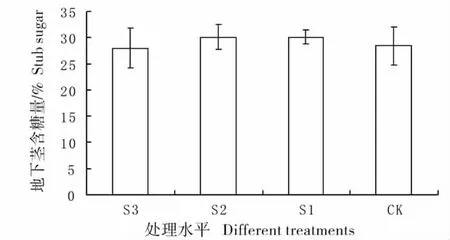
图7 不同遮荫处理下羊草地下茎的含糖量Fig.7 Percentage content of sugar of stub under different shading treatments
2.6 不同遮荫处理下羊草地下茎的含糖量变化
羊草地下茎含糖量较高,结合表1,可以得出,地下茎是重要的能量储藏器官,大约占整个植株能量的1/4 左右。各处理间含糖量并无显著差异(图7),最高值是S2 处理,含糖量达到30%左右,最低的是CK 组,含糖量在25%左右,说明遮荫并没有显著改变羊草地下茎的含糖量。
2.7 不同遮荫处理下羊草根茎的含糖量变化
几种处理间糖含量存在显著差异(图8),S1 处理的糖含量最高并与CK 和S3 处理达到差异显著水平,说明在轻度遮荫情况下,在实验收获时期羊草的根茎含糖量最高。
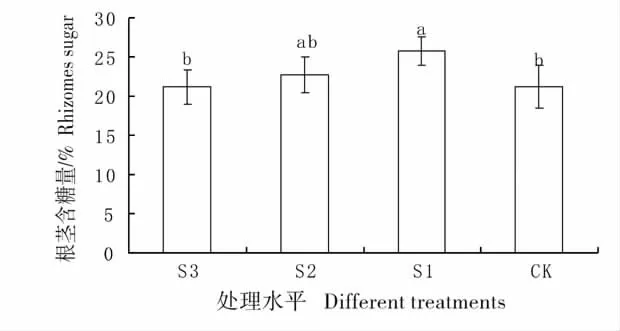
图8 不同遮荫处理下羊草根茎的含糖量Fig.8 Percentage content of sugar of rhizomes under different shading treatments
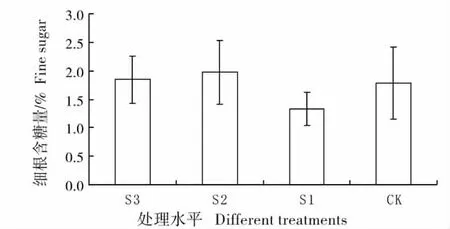
图9 不同遮荫处理下羊草细根的含糖量Fig.9 Percentage content of sugar of fine root under different shading treatments
2.8 不同遮荫处理下羊草细根的含糖量变化
图9 为不同遮荫处理下羊草细根含糖量变化,细根是羊草含糖量最少的部分,含糖量最多不超过2%,并且各种处理间无显著差异。S1 处理在其他器官内含糖量均较高,但细根含糖量确很低,说明羊草的光合产物并不是等量分配的,其能量投资利用具有不同的倾向性。
3 结论与讨论
实验结果表明遮荫干扰能够改变羊草分株种群光合产物的积累和分配。重度遮荫(70%)和轻度遮荫(30%)都出现了不同的能量分配和生殖投资等倾向性,各处理间不同器官的能量分配也发生了一定变化。
重度遮荫(70%)处理,羊草地上、地下各器官间糖分含量基本都低于对照和其它处理,只有穗含糖量略高于其他处理;重度遮荫情况下羊草生物量也最小。说明重度遮荫影响了羊草植株的光合作用,造成碳水化合物合成减少,不利于植株正常生长,这与王仁忠等[14]研究结果基本一致。但穗含糖量增加,可能与重度遮荫情况下,羊草为了完成自身繁衍,而进行的一种极端投资有关。笔者先前研究了遮荫干扰下羊草的抽穗率变化情况,结果与本实验基本一致,70%遮荫情况下能显著提高羊草的抽穗率[15]。羊草自然情况下抽穗率很低,种子产量较少[16],这种优质牧草因为种子产量低,限制了跨地域性大面积人工种植,如果能在进一步实践中研究遮荫与羊草抽穗率之间关系和机理,以及遮荫情况下种子成熟度和发芽率等,将有可能解决羊草种子繁殖问题。
轻度遮荫(30%)处理各器官含糖量和地上生物量均较高,这与植物在高光强下发生“午休”现象有关[17],轻度遮荫能够减少“午休”现象时间或不发生午休现象,增加了光合生产时间,另外,遮荫也有利于减少土壤水分蒸发,使土壤相对湿度大于对照组,也能够促进光合作用,进一步提高了轻度遮荫的光合作用效率。利用轻度遮荫能够提高羊草植株的含糖量特点,可以应用到生产中,用以提高羊草品质和羊草产量。
天然羊草分株种群地下根茎大部分是几株相连的,这种克隆植物的典型特征在接受外来干扰时,会表现出一种互惠共生模式或繁殖投资倾向策略,表现在器官上,会有部分器官形态或养分物质含量发生变化[14]。本研究发现,地上茎、叶鞘、叶片、地下茎各处理间含糖量有一定差异,甚至达到极显著水平,说明养分在种群分株内进行了重新分配,可能不同器官其功能分工不同,在外来干扰下,为了完成不同的生活目的,器官间或植株间进行了二次能量分配。
[1]杨允菲,张宝田.不同阶段的光温因子对羊草种群种子生产性状的影响[J].草业科学,1991,13(5):8 -13.
[2]杜占池,杨宗贵.羊草呼吸作用与温度光照水分的关系[J].植物生态学与地植物学,1993,17(4):339 -344.
[3]王仁忠.放牧影响下羊草种群生物量形成动态的研究[J].应用生态学报,1997,8(5):505 -509.
[4]Callaway R M,Delucia F H. Biomass allocation of mountain and desert ponerosa pine:an analog for response to climate change[J].Ecology,1994,75(5):1474 -1481.
[5]Benson E J,Hartnett D C.The role of seed and vegetative reproduction in plant recruitment and demography in tall grass prairie[J].Plant Ecology,2006,87:163 -178.
[6]董如磊,俞方圆,欧阳献.遮荫对东京野茉莉幼苗叶片形态和解剖结构的影响[J].江西农业大学学报,2010,32(5):974 -981.
[7]刘贤赵,康绍钟.番茄不同生育阶段遮荫对光合作用与产量的影响[J].园艺学报,2002,29(5):427 -432.
[8]刘广林,林瑛,王策.不同遮荫处理对几种扶芳藤光合特性影响的研究[J].园艺学报,2010,12(19):78 -81.
[9]杨持,杨理.光强梯度对羊草无性系分化与生长的影响[J].应用生态学报,1997,8(1):82 -87.
[10]Christa P H. Relationships between size,biomass allocation,reproduction,and survival in Triglochin palustris:implications for the effects of goose herbivory[J].Can J Bol,1998,76:2164 -2176.
[11]王德利,祝廷成.不同种群密度下羊草生态场梯度及其变化特征[J].应用生态学报,1995,6(1):1 -6.
[12]李海燕,李建东,徐振国,等.内蒙古图牧吉自然保护区羊草种群营养繁殖特性的比较[J].草业学报,2011,20(5):19 -25.
[13]王洪义,王正文,陈雅君.羊草分株种群特征及其繁殖策略对遮荫干扰的响应[D].哈尔滨:东北农业大学,2005.
[14]王仁忠.羊草种群能量生殖分配的研究[J].应用生态学报,2000,11(4):591 -594.
[15]王洪义,王正文,李凌浩,等.不同生境中克隆植物的繁殖倾向[J].生态学杂志,2005,24(6):670 -676.
[16]郭继勋,祝廷成.气候因子对东北羊草草原群落产量影响分析[J].植物学报,1994,36(10):790 -794.
[17]Harmony J D,David C H.The effects of fire frequency and grazing on tall grass prairie productivity and plant composition are mediated through bud bank demography[J].Plant Ecology,2009,201(2):411 -420.
