具有阶段结构的捕食-食饵模型的定性分析
2013-10-29姚若飞李艳玲
姚若飞,李艳玲
(陕西师范大学 数学与信息科学学院,陕西 西安710062)
文献[1-3]讨论了捕食者具有阶段性结构的捕食与被捕食模型
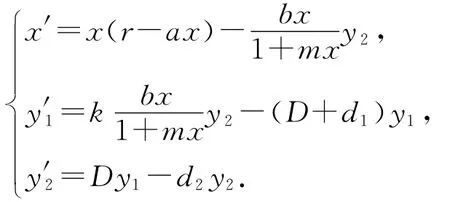
其中x(t)、y1(t)、y2(t)分别表示在时刻t食饵的种群密度、不成熟的和成熟的天敌种群密度.他们讨论了该常微分系统的一致持久性、正平衡态的存在性,及一定条件下的全局渐近稳定性和周期解的存在性与稳定性.
考虑到实际问题中种群密度与地域具有一定关系,因此本文讨论如下的反应扩散模型:
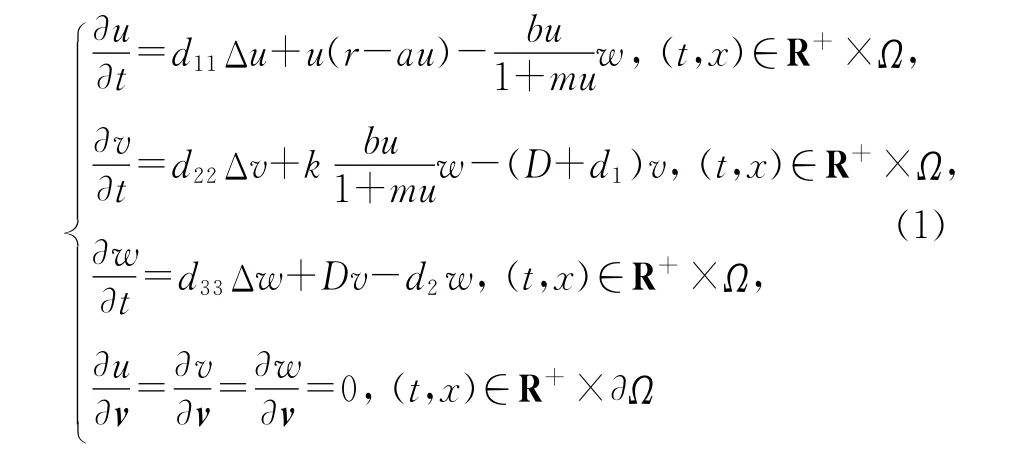
及初值条件
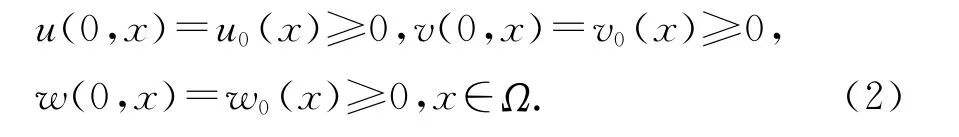
其中u0、v0、w0∈C2(Ω)∩C(¯Ω).这里Ω是Rn中的一个光滑区域,u(t,x)、v(t,x)、w(t,x)分别表示在时刻t、位置x处食饵的种群密度、不成熟的和成熟的天敌种群密度,v表示边界∂Ω上的单位外法向量,d11、d22、d33>0是扩散系数,r、a、b、m、k、D、d1、d2都为正常数.
为方便讨论稳态解问题,记d11=d22=d33=d,这实际上也是各个参数的一个转化,于是得到
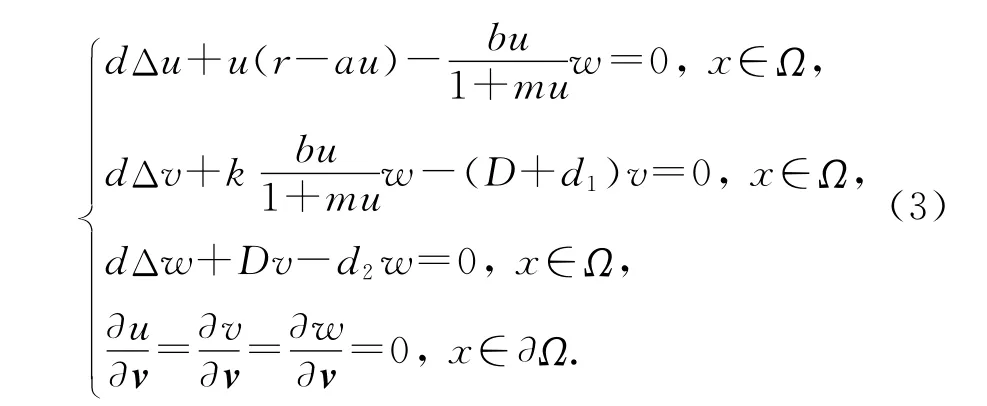
容易计算,系统(1)有一个平凡的常数稳态解U0(0,0,0)和一个半平凡的常数稳态解U1(r/a,0,0),另外当满足条件时,系统(1)还有一个正的常数稳态解U*(u*,v*,w*),这里

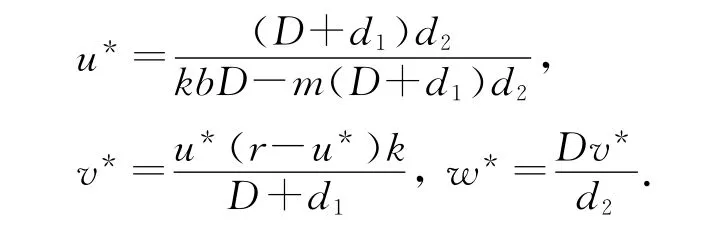
1 整体解的存在性和全局稳定性
初边值问题(1)-(2)的解是局部存在唯一的[4],这里只讨论整体解的存在性.注意到两类捕食者之间是互惠关系[5],因此对v、w的方程,方程组比较原理是成立的,于是有如下两个结论:
定理1 设(u,v,w)∈C2([0,T)ׯΩ,R3)是系统(1)-(2)的一个解,那么(u,v,w)是非负的,并且有一个至多是T的指数函数的上界.
定理2 设d2(D+d1)(a+mr)>kbrD.如果(u,v,w)∈C2([0,∞)ׯΩ,R3)是系统(1)-(2)的一个正解,则对x一致地有

定理1和定理2的证明 易知[0,+∞)3是系统(1)的一个不变区域,故(u,v,w)非负,并且当且仅当u0≢0时,u>0,∀t>0,x∈¯Ω;当且仅当v0≢0或w0≢0时,v、w>0,∀t>0,x∈¯Ω.下面讨论正解情形,设U(t)是常微分系统
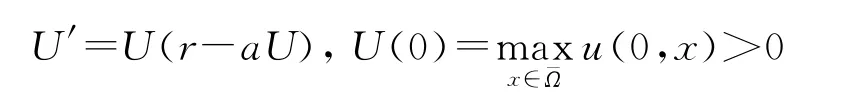
的解,那么当t→+∞时,U(t)→r/a.对比u的方程,利用单个方程的比较原理知

从而u有界.注意到bu/(1+mu)<b/m,令(V(t)、W(t))是线性常微分系统

的解,于是(V(t),W(t))至多指数增长.因为(v,w)的方程是互惠的,由方程组的比较原理得
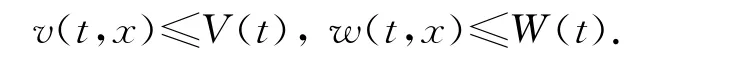
所以,(u,v,w)有界并且有一个至多是T的指数函数的上界,从而系统(1)-(2)的解是整体存在的.
在定理2的条件下,存在R>r,使d2(D+d1)(a+mR)>kbRD.并且对x一致地有
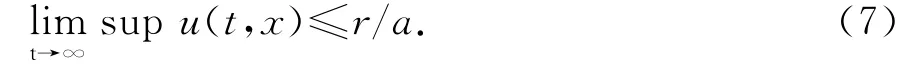
取T1>0使得当t≥T1时,u(t,x)≤R/a.构造(V(t),W(t))如(6),初值为T1时刻代替,则由R的取法知(V(t),W(t))指数收敛于(0,0).从而由比较原理可知,v(t,x)≤V(t),w(t,x)≤W(t),t≥T1,x∈¯Ω.于是
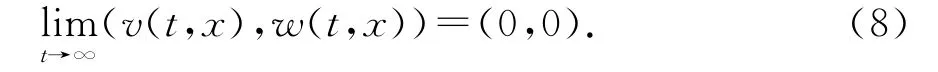
对任意的ε∈(0,r/ab),取T2>T1,使得当t≥T2时v(t,x)<aε.令u~(t)如下:

因为bwu/(1+mu)≥abεu(当t≥T2时),利用u的方程及比较原理,得
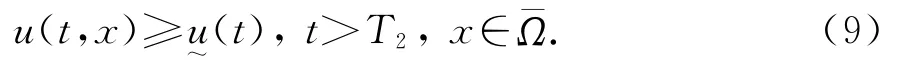
注意到u~(t)→r/a-bε,及(7)-(9)和ε的任意性知定理2的结论成立.
2 正解的先验估计和非常数正解的不存在性
本节应用一个与最大值原理有关的引理,讨论正解的估计,利用这个估计和能量积分方法得到一个非常数正解的不存在性结果.
引理1[6]设F(x,w)∈C(¯Ω×R1).若w∈C2(Ω)∩C1(¯Ω)满足
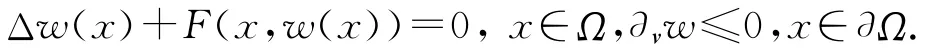
如果w(x0,那么F(x0,w(x0))≥0.
定理3 设(u,v,w)是系统(3)的一个正解,则0<u<r/a,0<v<K,0<w<K,x∈¯Ω.
其中K是一个与m、b、d无关的正常数.
令V=ku+v,则它满足

于是得到

定理4 存在一个仅依赖r、a、b、k、D、d1、d2、Ω的正常数D,使得当d>D时,系统(3)的正解一定为正常数.
证明 若φ∈L1(Ω),记ˉφ=∫Ωφ(x)dx/|Ω|和φ=φ-ˉφ.令n(u)=u(r-au),f(u,w)=buw/(1+mu).如果(u,v,w)是系统(3)的一个正解.对(3)中3个方程分别乘以u、v、w后,再在Ω 上积分,利用Green公式,得
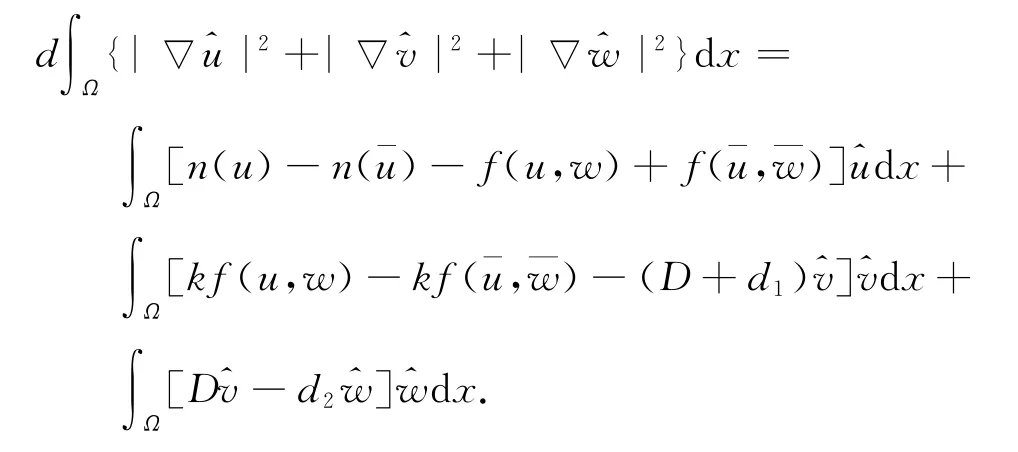
注意到n(u)-n(u)=g1u,f(u,w)+f(ˉu,ˉw)=g2u+g3w,这里g1、g2、g3都是函数且有不依赖于m和d的上界(已用到定理3),那么

结合Poincar˙e不等式
∫Ω|φ-ˉφ|2dx≤CΩ∫Ω|▽φ|2dx,∀φ∈W1,2(Ω)和Young不等式,得


上式表明,当d>CΩK1(K1不依赖于m和d)时,u=v=w=0,即u、v、w都是常函数.
3 常数稳态解处的局部分歧
本节讨论系统(3)在3个常数平衡态的局部分歧,其主要是验证局部分歧定理(文献[8]中定理13.5)中的3个条件成立.
设0=λ0<λ1≤λ2≤…→+∞是-Δψ=λψ,x∈Ω,∂vψ=0,x∈∂Ω (10)的所有特征值,相对应的单位正交化的特征函数依次为ψ0,ψ1,ψ2,….ψ0是主特征函数,为正函数,其他特征函数均变号.
Hilbert空间[L2(Ω)]3具有内积〈U,V〉=,U=(u1,u2,u3)T,V=(v1,v2,v3)T,以后讨论中出现垂直(符号为⊥)的概念就是在这种意义下的.记
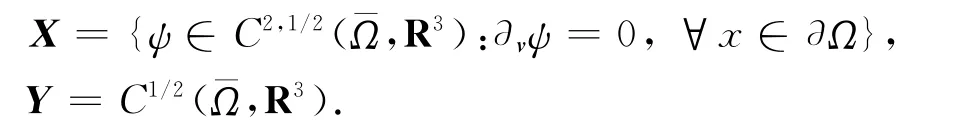
它们在通常的范数意义下都是Banach空间.设U=(u,v,w),
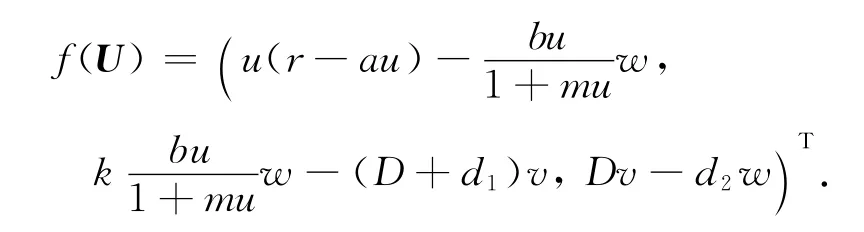
定义光滑算子F:R+×X→Y为F(d,U)=dΔU+f(U).那么系统(3)转化成

则FU(d,U)=dIΔ+H(U):X→Y,FdU(d,U)=IΔ:X→Y,其中

先计算算子L=IΔ+H:X→Y的一些性质(H是实矩阵,HT表示H的转置).设U∈Ker(L),并记于是有
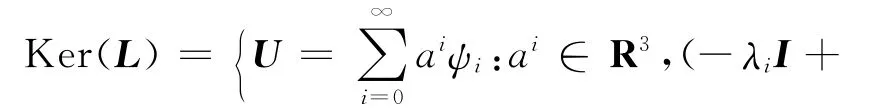
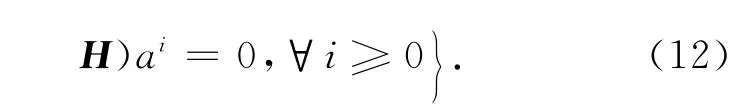
上述的求和实际上是有限和,而R(L)={f∈Y:f⊥Ker(L*)},L*=IΔ+HT,故

由此,Ker(L)∩R(L)= {0}的充要条件是对每一个i,λi(关于矩阵H)的代数重数和几何重数相等(可能都为0).这里(12)式用来计算L的核空间,(13)式用来判断L的像空间的余维数,而(14)式是在以后的情形中用来判断分歧定理第三个条件的.
在高维空间中,特征问题(10)的正特征值的重数可能大于1.然而在一维区间时,所有特征值都是单重的,这里讨论空间为一维的情形:

对平衡态U0(0,0,0)和U1(r/a,0,0),F(d,U0)=F(d,U1)=0,∀d>0.

显然,H0的正特征值只有一个,即为r,并且代数重数是1,以及0不是特征值;当d2(D+d1)(a+mr)<kbrD时,H1的正特征值只有一个,记为σ,并且代数重数是1,以及0不是特征值.对于这两种平衡态,其分歧情形是相似的.
定理5 给定j≥1和d=r/λj,则存在δ>0和光滑曲线(d,φ):(-δ,δ)→R×Z满足:(ⅰ)φ(0)=0,d(0)=d,(ⅱ)F(d(s),U(s))=0,s∈ (-δ,δ);(ⅲ)在(d,U0)的某个小邻域内(11)的每个解要么在这条曲线上,要么是平凡的.这里Φ=ξψj,Z={φ∈X:φ⊥Φ},ξ= (1,0,0)T是H0的正特征值所对应的“唯一”特征向量及
U(s)=U0+s(Φ+φ(s)).
证明 设L0=FU(d,U0)=+H0,L1=IΔ,则由(12)得,Ker(L0)=span{Φ},Φ =ξψj,L1Φ=-λjΦ.再由(13)和(14)式得,codimR(L0)=1,L1Φ =-λjΦ ∉R(L0).分歧定理的条件成立.
定理6 设条件(4)成立,且记σ是H1上的唯一正特征值.给定j≥1和d=σ/λj,则存在δ>0和光滑曲线(d,φ):(-δ,δ)→R×Z满足:(ⅰ)φ(0)=0,d(0)=d;(ⅱ)F(d(s),U(s))=0,s∈(-δ,δ);(ⅲ)在(^d,U1)的某个小邻域内系统(11)的每个解要么在这条曲线上,要么是平凡的.这里Φ=ξψj,Φ=ξψj,Z={φ∈X:φ⊥Φ},ξ、ξ是H1和HT1的正特征值所对应的“唯一”特征向量及

下面讨论正常数平衡态U*处的分歧,记

那么矩阵H*的特征多项式是

注意到r-au*>0,那么H*至少有一个负特征值,0不是特征值,并且它的正特征值(若有的话)必然是两个.假设
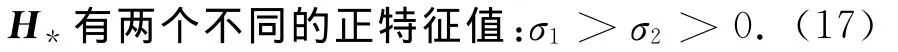
为避免σ1/λj=σ2/λi的情形,进一步设

同样地,有下述分歧结论:
定理7 设条件(4)、(17)、(18)成立.给定i∈{1,2},j≥1.记d=σi/λj,则存在δ>0和光滑曲线(d,φ):(-δ,δ)→R×Z满足:(ⅰ)φ(0)=0,d(0)=d;(ⅱ)F(d(s),U(s))=0,s∈(-δ,δ);(ⅲ)在(^d,U*)的某个小邻域内(11)的每个解要么在这条曲线上,要么是平凡的.这里Φ=ξψj,Φ=ξψj,Z= {φ∈X:φ⊥Φ},ξ、ξ是H*和HT*的正特征值σi所对应的“唯一”特征向量及
U(s)=U*+s(Φ+φ(s)).
证明 同前面定理证明一样,这里只需要验证单特征值分歧定理的第三个条件,事实上,σi(i=1,2)是H*的代数单重特征值就表明ξ·ξ≠0.从而结论得证.
注1 取参数值b=1、m=1、D=1、d1=0.1、d2=0.2、k=1、a=2、r=12时,U*=(0.282 1,2.932 3,14.661 4),H*的特征值为-1.634 7,1.469 7,0.816 8.这表明条件(17)在某些参数的时候是可以成立的,而条件(18)只要合适地扰动参数值也可以成立.
[1]Wang Wendi.Global dynamics of a population model with stage structure for predator[C]∥Chen Lansun.Advanced Topics in Biomathematics.River Edge:World Scientific Publishing,1998:253-257.
[2]Xiao Yanni,Chen Lansun.Global stability of a predator-prey system with stage structure for the predator[J].Acta Mathematica Sinica,2004,20(1):63-70.
[3]Georgescu Paul,Hsieh Ying-Hen.Global dynamics of a predator-prey model with stage structure for the predator[J].SIAM Journal on Applied Mathematics,2007,67(5):1379-1395.
[4]Avner Friedman.Partial differential equations of parabolic type[M].Prentice-Hall:Englewood Cliffs,1964.
[5]叶其孝,李正元.反应扩散方程引论[M].北京:科学出版社,1990:192-235.
[6]Lou Yuan,Ni Weiming.Diffusion,self-diffusion and cross-diffusion[J].Journal of Differential Equations,1996,131(1):79-131.
[7]Jang Jaeduck,Ni Weiming,Tang Moxun.Global bifurcation and structure of turing patterns in the 1-D Lengyel-Epstein model[J].Journal of Dynamics and Differential Equations,2004,16(2):297-320.
[8]Smoller J.Shock waves and reaction-diffusion equations[M].New York:Springer-Verlag,1994:167-191.
[9]马翠,李艳玲.一般的Gause型捕食-食饵模型的定性分析[J].陕西师范大学学报:自然科学版,2010,38(3):6-9.
[10]Wu Jianhua.Global bifurcation of coexistence state for the competition model in the chemostat[J].Nonlinear Analysis,2000,39(7):817-835.
