黑鳃梅童鱼和棘头梅童鱼的形态学比较研究*
2013-10-16高天翔马国强
高天翔,韩 刚,2,马国强,宋 娜
(1.中国海洋大学海洋生物多样性与进化研究所,山东 青岛266003;2.中国水产科学研究院,北京100141)
黑鳃梅童鱼(Collichthys niveatus)与棘头梅童鱼(Collichthys lucidus)同属于鲈形目(Perciformes)、石首鱼科(Sciaenidae)、梅童鱼属(Collichthys),均为我国重要的小型底栖经济鱼类[1]。黑鳃梅童鱼常见于黄渤海;棘头梅童鱼广泛分布于黄渤海、东海和南海[1-2]。2种梅童鱼适温、适盐广,均为沿海居民喜食的鱼类。每年的4~6月和9~10月为其渔汛盛期。国内有一些梅童鱼相关研究报道,但多为其资源、生物学特性及遗传学等方面的研究[3-5]。迄今尚未见关于棘头梅童鱼与黑鳃梅童鱼形态学比较研究报道。
形态学研究方法,又称生物学测定法,是从表型性状上研究物种分类的最简便、最古老的方法,属于传统的鱼类系统发育学研究方法。迄今,形态学方法仍是鱼类分类研究的主要手段,通过对形态可量、可数性状的观察和统计学分析,可以在短时间内将所研究物种归类,并对物种的遗传变异有所了解。以前,黑鳃梅童鱼与棘头梅童鱼主要依据外部形态特征的差异(鳔侧枝数目、鳃腔颜色等)进行分类。近年来,部分遗传学研究结果与以往形态学结论不同,因此对这2个物种的划分存在疑问[6]。为此,本研究将黑鳃梅童鱼与棘头梅童鱼的形态差异进一步明晰,并将传统形态学度量方法和统计学比较分析相结合,阐明2种梅童鱼间形态学差异,以期为石首鱼类种类鉴别、为梅童鱼分类研究提供基础资料。
1 材料与方法
1.1 材料

表1 黑鳃梅童鱼和棘头梅童鱼的采样信息Table 1 The information of sampling locations,sampling dates and sample sizes for C.niveatus and C.lucidus
1.2 实验方法
1.2.1 形态拍照 对2种梅童鱼顶枕部中央小棘、臀鳍第一鳍棘、鳃腔、鳔及耳石进行拍照,比较分析2种梅童鱼的形态差异。
1.2.2 传统形态学数据测量方法 本研究采用传统的形态学测量方法,对样本的10个可数性状进行计数(背鳍鳍条数、胸鳍鳍条数、尾鳍鳍条数和臀鳍鳍条数以及脊椎骨数、上鳃耙数、下鳃耙数、鳔侧枝数)。并对16个可量性状(体长、体高、尾柄长、尾柄高、头长、吻长、上颌长、眼径、眼间距、眼后头长、吻至背鳍起点、腹鳍起点至臀鳍起点、背鳍基长、臀鳍基长、腹鳍长、胸鳍长)用游标卡尺进行测量,长度精确到0.01mm,各测量指标如图1所示。用电子天平测量体重,重量精确到0.1g。

图1 黒鳃梅童鱼和棘头梅童鱼的16个形态学测量指标Fig.1 The 16morphometric characters for C.niveatus and C.lucidus
1.2.3 数据处理 将16个传统形态学数据进行标准化后,得到14组比例性状变量,用统计软件SPSS 11.5,分别对其进行主成分分析、判别分析和单因子方差分析。
A~N依次表示14组标准化后的形态学数据比值:A体高/体长、B尾柄长/尾柄高、C头长/体长、D吻长/头长、E上颌长/头长、F眼径/头长、G眼间距/头长、H眼后头长/头长、I吻至背鳍起点/体长、J腹鳍起点至臀鳍起点/体长、K背鳍基长/体长、L臀鳍基长/体长、M腹鳍长/体长、N胸鳍长/体长。
1.2.3.1 判别分析 利用对2种梅童鱼形态差异贡献率较大的参数建立判别公式,判别准确率按下式计算:
判别准确率P(%)=(判别正确的个体数/实际个体数)×100%;
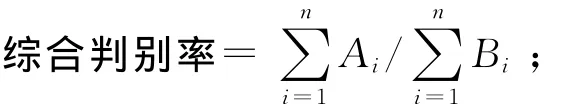
式中:实际个体数为该种群的样本数,判别个体数为被判别为该种群的个体数;Ai为第i个种群中判别正确的个体数;Bi为第i个种群的实际个体数;n为种群数。
1.2.3.2 主成分分析 运用 SPSS 11.5软件,分别对黑鳃梅童鱼和棘头梅童鱼的形态学数据进行主成分分析,得到各主成分的贡献率和特征值,并根据主成分得分绘制主成分散布图。
马来西亚橡胶木已于2017年7月1日起全面禁止出口,这对缓解相关产业的原料短缺问题,尤其是对家具制造业有所帮助。到2020年马来西亚家具业对出口额实现120亿林吉特的目标也更充满了信心。
1.2.3.3 单因子方差分析 运用SPSS 11.5软件的单因子方差分析(one-way ANOVA)分析2种梅童鱼形态学差异。
2 结果
2.1 黑鳃梅童鱼与棘头梅童鱼形态学特征差异
对2种梅童鱼及其相应部位进行拍照(见图2)。黑鳃梅童鱼顶枕部的中央有2个小棘,呈镰刀状。臀鳍第一鳍棘略弯曲,呈钩状。鳔细长,达腹腔的前后端,两侧约有13~16对树枝状侧枝。鳃腔皮肤及鳃耙具有黑色素沉积。耳石较厚,表面及边缘较平滑。棘头梅童鱼顶枕部的中央有一列小棘,约4~6个。臀鳍第一鳍棘粗壮尖直,约为第二鳍棘的2/5。鳔两侧树枝状分支较黑鳃梅童鱼更密,约20~24对。鳃腔和鳃耙均为鲜红色。耳石相对较薄,整体比黑鳃梅童鱼小,表面凹凸不平,存在较多瘤状突起,边缘存在锯齿状突起。
2.2 形态学数据分析
2.2.1 分节特征 表2显示了2种梅童鱼形态学可数性状的测量结果。从表2中可以看出,2种梅童鱼的背鳍鳍条数、胸鳍鳍条数、尾鳍鳍条数、臀鳍鳍条数、脊椎骨数、上鳃耙数、下鳃耙数平均值分别为26.66和26.50,13.69 和 14.77,18.97 和 20.57,12.35 和11.63,25.97和27.20,8.21和9.57,17.35和16.80;鳔侧枝数差异明显,平均值分别为14.35和22.07。

表2 黑鳃梅童鱼和棘头梅童鱼可数性状Table 2 Meristic counts of C.niveatus and C.lucidus
2.2.2 判别分析 为减少个体实际长度大小对可量性状的影响,利用SPSS软件中的相关分析对每条鱼的可量性状进行两两组合求其相关性。以其中的14组数据作为标准化值进行判别分析。为了确保判别准确性,利用贡献率大的8个参数建立2种梅童鱼的判别公式。

图2 黑鳃梅童鱼与棘头梅童鱼形态差异Fig.2 The morphological difference between C.niveatus and C.lucidus
黑鳃梅童鱼:

棘头梅童鱼:

式中:X1~X8分别代表:B尾柄长/尾柄高、E上颌长/头长、F眼径/头长、G眼间距/头长、H眼后头长/头长、L臀鳍基长/体长、M 腹鳍长/体长、N胸鳍长/体长。将校正过的形态参数(X1~X8)代入上述判别公式中,即可判断该鱼所属种,判别准确率为100%。2.2.3主成分分析 对2种梅童鱼的可量性状进行主成分分析,分析结果见表3。从表3中可以看出,前6项主成分的累计贡献率达到78.147%,即前6个主成分可以解释2种梅童鱼间形态差异的78.147%。其中第1到第6主成分贡献率分别为20.328%、17.931%、12.053%、11.365%、9.116%、7.354%。从表3可以看出,对第1主成分载荷值较大的是B(尾柄长/尾柄高)、C(头长/体长)、F(眼径/头长);对第2主成分载荷值较大的是A(体高/体长)、E(上颌长/头长)、H(眼后头长/头长);对第3主成分载荷值较大的是J(腹鳍起点至臀鳍起点/体长)、K(背鳍基长/体长)、L(臀鳍基长/体长);对第4主成分载荷值较大的是M(腹鳍长/体长)、N(胸鳍长/体长);对第5主成分载荷值较大的是D(吻长/头长)、E(上颌长/头长)、L(臀鳍基长/体长);对第6主成分载荷值较大的是H(眼后头长/头长)、J(腹鳍起点至臀鳍起点/体长)、L(臀鳍基长/体长)。

表3 黑鳃梅童鱼和棘头梅童鱼可量性状的主成分分析结果Table 3 The principle compoment analysis of C.niveatus and C.lucidus
根据第1和第3主成分以及第2和第3主成分分别绘制散点图(见图3)可以看出,黑鳃梅童鱼和棘头梅童鱼在外部形态可量数据上虽存在个别交叉,但仍能形成各自相对集中的组。

图3 黑鳃梅童鱼与棘头梅童鱼第1和第3主成分以及第2和第3主成分分布图Fig.3 Plot of the 1st,3rdand 2nd,3rd principle components from PCA based on the morphological characteristics between C.niveatus and C.lucidus
2.2.4 单因子方差分析(one-way ANOVA) 对2种梅童鱼14个形态特征比值进行单因子方差分析,同时对各群体形态学指标进行多重比较分析,对具有方差齐性的变量采用LSD比较法分析,不具有方差齐性的变量采用Tamahane’s T2比较法进行分析。P=0.01显著性水平下的单因子方差分析结果如表4所示,黑鳃梅童鱼和棘头梅童鱼在7个变量(B,E,F,G,H,L,N)上存在显著性差异。
3 讨论
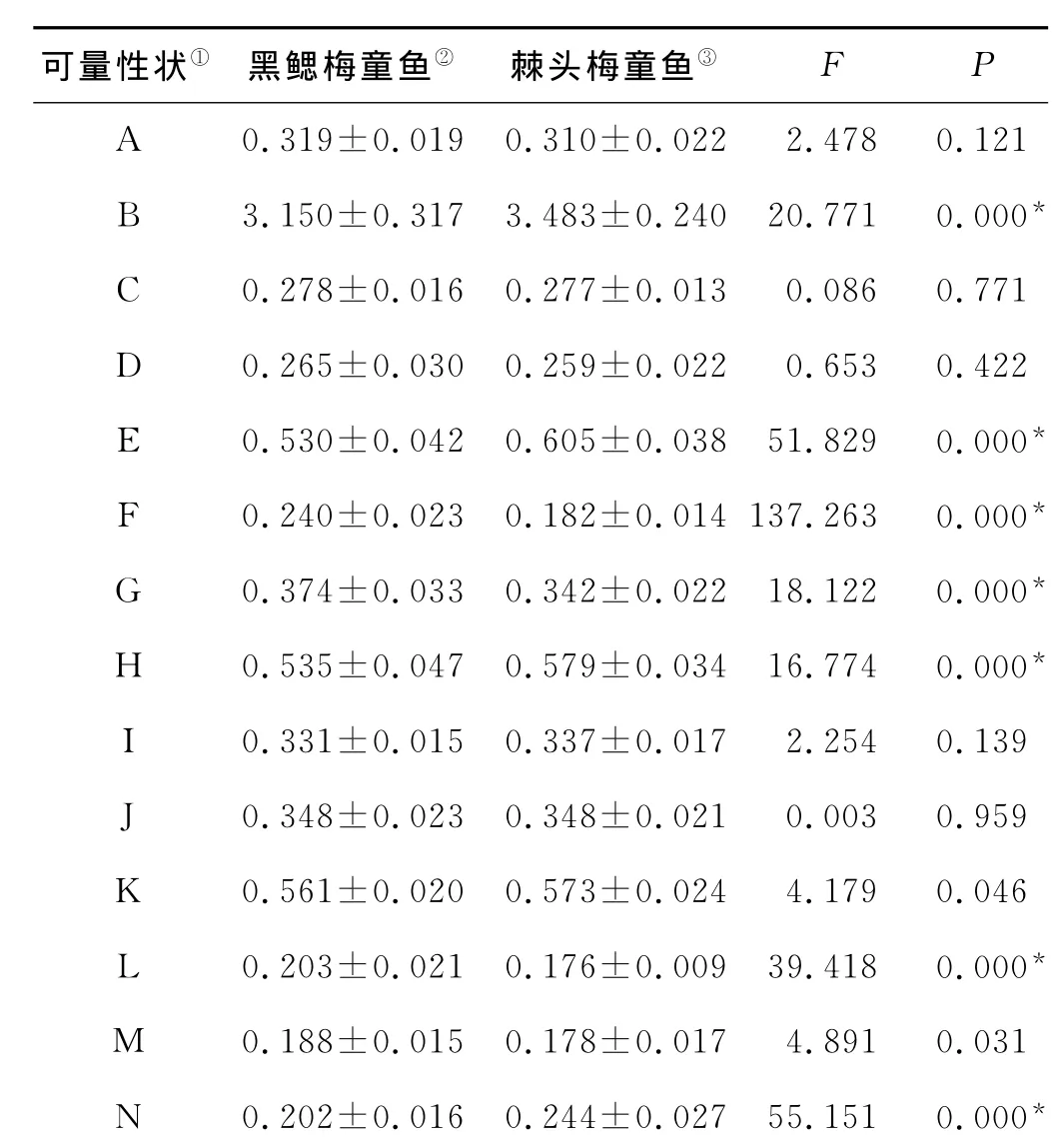
表4 黑鳃梅童鱼和棘头梅童鱼单因子方差分析结果(平均值±标准差,P=0.01)Table 4 The results of one-way ANOVA between C.niveatus and C.lucidus(Mean±S.D.,P=0.01)
3.1 黑鳃梅童鱼与棘头梅童鱼不同形态特征比较
从照片可以直观地看出,2种梅童鱼在顶枕部小棘、臀鳍第一鳍棘、鳔侧枝、鳃腔颜色和耳石等形态特征上存在明显不同(见图3),与朱元鼎等[1]的分类描述一致,利用上述部位应可对2种梅童鱼进行快速鉴定。
3.2 两种梅童鱼的分节特征
从可数性状的结果来看,2种梅童鱼的背鳍、胸鳍、尾鳍、臀鳍、脊椎骨数、上鳃耙数和下鳃耙数分别在23~28、11~17、18~21、11~13、25~28、6~11和15~19之间,与朱元鼎等[1]研究结果基本一致。2种梅童鱼鳔侧枝数有显著差异(黑鳃梅童鱼为13~16,棘头梅童鱼为20~24)。由于2种梅童鱼的鳍条数、脊椎骨数和上下鳃耙数均存在重叠,并无明显差异,因此分节特征上仅鳔侧枝可作为鉴别2种梅童鱼的有效指标。
3.3 判别分析、主成分分析和单因子方差分析
判别分析是根据若干变量来判断所研究对象如何分类,具有较强实用性,是被广泛应用于各种领域的统计方法[7-12]。目前判别分析中最常用的方法为逐步判别法,它从若干已知因子中挑选最佳的、必要的因子来建立判别函数。本研究采用逐步判别法从14个形态学数据中筛选出8个主要性状并建立判别公式,其判别准确率为100%,说明基于形态学数据的逐步判别法对2种梅童鱼的判别也是有效的。
主成分分析是1种多变量的分析方法,运用该方法能够较直观的评价个体之间的相似程度,已被广泛应用于鱼类不同群体间差异程度[13-14]和对新物种的发现[15-16]等问题的研究。主成分分析结果显示,2种梅童鱼的形态学数据在第1和第3主成分以及第2和第3主成分上除存在个别交叉现象外,均能够明显分为两大部分。
单因子方差分析(one-way ANOVA)是用来检验由单一因素影响的几个相互独立的组是否来自服从正态分布的总体。本研究中棘头梅童鱼和黑鳃梅童鱼群体均来自正态分布的总体,适合单因子方差分析。研究结果显示,P=0.01时,2种梅童鱼在形态学数据上有7个变量(B,E,F,G,H,L,N)存在显著差异。
传统形态学分析方法在鱼类的比较、分类研究中是最基本、最主要的研究方法之一。通过形态性状可以在短时间内将所研究物种归类,并对物种的遗传变异有所了解,因此,形态学方法是一种简单、经济、快速的方法[17]。本文从形态学角度采用多种分析手段,对2种梅童鱼进行了比较研究,结果表明2种梅童鱼在外部形态上存在明显差异。以上形态学研究结果与马国强等基于16SrRNA和Cyt b基因序列的遗传学研究结论相同[18]。由于梅童鱼幼鱼的形态与小黄鱼等石首鱼类极其相似,因此要慎重进行幼鱼种类的鉴定,必要时需要形态学和遗传学研究方法相结合。
[1] 朱元鼎,伍汉霖.东海鱼类志 [M].北京:科学出版社,1963:286-293.
[2] 杨纪明.渤海鱼类的食性和营养级研究 [J].现代渔业信息,2001,16:10-19.
[3] 吴振兴,吴常文.浙江近海棘头梅童鱼生长规律与群体组成的研究 [J].水产科技情报,1990,17:170-174.
[4] 谢起浪,单乐州,王铁杆,等.浙南飞云江口棘头梅童鱼的资源调查和研究 [J].渔业现代化,2006,1:35-36.
[5] 单乐州,邵鑫斌,谢起浪,等.棘头梅童鱼人工繁殖及育苗技术研究 [J].浙江海洋学院学报,2006,25:266-271.
[6] 蒙子宁,庄志猛,丁少雄,等.中国近海8种石首鱼类的线粒体16SrRNA基因序列变异及其分子系统进化 [J].自然科学进展,2004,14(5):514-521.
[7] 张尧庭,方开泰.多元统计分析引论 [M].北京:科学出版社,1982:237-247.
[8] 詹秉义.渔业资源评估 [M].北京:中国农业出版社,1995:20-21.
[9] 任一平,刘群,李庆怀,等.青岛近海小型鳀鲱鱼类渔业生物学特性的研究 [J].海洋湖沼通报,2002,1:69-74.
[10] 江胜峰,程家骅.海洋小型鱼类研究进展 [J].海洋渔业,2006,28:336-340.
[11] Ricker W E.Computations and Interpretation of biological statistics of fish populations [M].Canada:Bulletin of the Fisheries Research Board of Canada,1975,290-293.
[12] Nakabo T.Fishes of Japan with Pictorial Keys to the Species(Second edition)[M].Kanagawaken:Tokai University Press.2000:822-827.
[13] Gibson A R,Baker A J,Moeed P.Morphometric variation in introduced populations of the common myna(Acridotheres tristis):An application of the jackknife to principal component analysis[J].Systematic Zoology,1984,33:408-421.
[14] Voss R S,Marcus L F,Escalante P.Morphological evolution in muroid rodents I.Conservative patterns of craniometric covariance and their ontogenetic basis in the neotropical genus Zygodontomys[J].E-volution,1990,44:1568-1587.
[15] Eisenhour D J.Systematics of Macrhybopsis tetranema (Cypriniforms:Cyprinidae)[J].Copeia,1999,4:969-980.
[16] Golubtsov A S,Berendzen P B.Morphological evidence for the occurrence of two electric catfish (Malapterurus)species in the White Nile and Omo-Turkana systems(East Africa)[J].Journal of Fish Biology,1999,55:492-505.
[17] Schaal B A,Leverich W J,Rogstad S H.Comparison of methods for assessing genetic variation in plant conservation biology[M].∥Falk I D A,Holsinger K E eds..Genetics and Conservation of Rare Plants.New York:Oxford University Press,1991:123-134.
[18] 马国强,高天翔,孙典荣.基于16SrRNA和Cyt b基因序列探讨2种梅童鱼的遗传分化 [J].南方水产,2010,6(2):13-20.
