湖北石首麋鹿昼间活动时间分配
2013-09-11杨道德李竹云李鹏飞蒋志刚
杨道德 ,李竹云,李鹏飞,蒋志刚
(1.中南林业科技大学野生动植物保护研究所,长沙 410004;2.湖北石首麋鹿国家级自然保护区管理处,石首 434401;3.中国科学院动物研究所,北京 100101)
动物必须合理地分配各种行为占用的时间才能生存繁衍[1]。在不同的生境中,或随着自身生理状态的变化,动物行为收支代价也随之改变,动物需尽快调整活动时间分配来适应这种变化[2]。通过对动物昼间活动时间分配的研究,可探讨性别、年龄和季节变化对动物行为的影响及其行为适应。麋鹿(Elaphurus davidianus)为中国特有种,曾在野外灭绝。我国从1985年开始重引入麋鹿,至今已发展到2000多头,并已在江苏大丰、湖北石首、北京麋鹿苑、河南原阳建立了4个较大的麋鹿圈养、散放或自然野化种群[3]。国外学者对圈养麋鹿种群的繁殖[4]、生态[5]等方面进行了研究;国内学者对麋鹿种群的发情期昼间活动时间分配[6-7]、行为谱[8]、种群动态[9-10]、濒危原因[11]、交配计策[12-13]、遗传多样性[14]、生境与行为[15]、栖息地退化[16]、记忆[17]、灭绝风险[18]、潜在生境评价[19]等方面已有报道,尚未见散放麋鹿种群全年昼间活动时间分配的研究报道,而这方面的研究结果对今后麋鹿的迁地保护、自然野化种群恢复和行为适应研究有重要意义。
1 自然概况
湖北石首麋鹿国家级自然保护区(以下简称石首麋鹿保护区),地处湖北省石首市天鹅洲长江故道西南端,南临长江防洪堤,东抵天鹅洲长江故道,中心地理坐标为112°33'E,29°49'N,围栏总面积约11.67 km2。石首麋鹿保护区地势低平,最高海拔38.44 m,最低海拔32.91 m。天鹅洲长江故道水面宽约1 500 m,一般水深4 m,在每年的汛期(5—9月份)与长江相通,枯水期(10月—翌年4月份)与长江隔绝。故道水体pH值为7.00—7.49,水质达中等软水标准。石首麋鹿保护区属亚热带季风气候区,夏季炎热,冬季干冷。年均气温16.5℃,最热月(7月份)平均气温28.5℃,最冷月(1月份)平均气温3.5℃。年均相对湿度约80%。年均降水量1 200 mm以上。无霜期长达26l d。植被类型可分为8种主要群落[10]。
2 研究方法
2.1 季节划分
石首麋鹿保护区地处中亚热带北部,属典型的亚热带季风湿润气候区,春、夏、秋、冬四季的划分时段分别为3—5月、6—8月、9—10月、11月—翌年2月。
2.2 研究对象与取样
成年麋鹿指有繁殖能力且体成熟的个体;亚成年麋鹿指处于生长发育的个体[8]。根据体形、尾长、后足长、角的有无及分支、年龄等综合特征[8-9],本研究将观察对象分为4个性别-年龄组:成年雄鹿(3岁以上)、成年雌鹿(2岁以上)、亚成年鹿(1—3岁雄鹿和1—2岁雌鹿)、幼鹿(1岁以下)。麋鹿的行为可分为摄食、休息、运动、排遗、调温、发情、交配、分娩、育幼、冲突、通讯、聚群、杂类等13大类型[9]或采食、饮水、排泄、运动、休息、集群、自卫防御、繁殖、信息传递、智能等10大类型[8]。到2006年12月底时,石首麋鹿保护区围栏内的麋鹿种群已达522头。为了方便观察和记录,本研究将麋鹿个体昼间活动行为归为6大类型:(1)采食:采食草、嫩叶、芦苇嫩茎,刨食草根、幼鹿吸乳等;(2)饮水:寻找水源、饮水;(3)运动:散步、奔跑、跳跃拍打、角斗等活动;(4)休息:侧躺、站、犬坐,低头闭目睡眠而四肢不动,仅头部观望或口中反刍食物,母鹿哺乳;(5)警觉:单个或群体对外界变化所做出的反应,如警戒和自我修饰、通迅、背腹涂泥浆、浸于水中、甩尾驱虫等活动;(6)其他行为:上述5种行为以外的其他行为,如冲突、排泄、交配等。
行为取样采用焦点取样法[20]:观察时随机选定某一个体作为观察对象进行连续观察,直到观察对象从视野中消失为止。记录采用瞬时取样法:每次取样间隔1min,记录所观察个体的性别、年龄、活动时间、空间位置及行为类型。2006年9月—2007年10月,每月对麋鹿昼间行为跟踪观察6—8 d(双筒望远镜,Steiner 1 0.5×28),每个性别1年龄组的观测样本量为20个个体。夏、秋季跟踪观察时间为6:00—20:00,冬、春季为7:00—19:00;观察距离为80—150 m。为便于不同季节间的比较[21],将不同季节每天7:00—19:00的观察时间段按2 h等分为6个时段。有效工作时间总计114 d,共计427.5 h(其中:春季98.7 h、夏季142.9 h、秋季83.6 h、冬季102.3 h),总记录数据15 230个。文中数据为平均数±方差,除特殊说明外。
2.3 数据处理与分析
首先对每天记录的各时段各种行为类型的时间进行统计,计算出所占百分比,再将每天各种行为类型的时间分别占当天总观察时间的百分率作为该日的活动时间分配。所有数据处理和统计分析用Excel 2003和SPSS 15.0软件完成。由于记录的全年行为数据经Kolmogorov-Smimov检验均呈非正态分布,故不同季节间的昼间活动时间分配的差异采用k个独立样本的非参数Kruskal Wallis H检验[22-23];不同性别-年龄组的昼间活动时间分配的差异采用两个独立样本的Mann-Whitney U-test进行统计分析[24]。不同季节各性别、年龄组的行为数据经Kolmogorov-Smimov检验均正态分布,故不同性别、年龄组在不同季节的昼间活动时间分配的差异采用方差(ANOVA)分析。显著水平设定为α =0.05。
3 研究结果
3.1 年均昼间活动时间分配
统计所观察的麋鹿个体的全年昼间活动时间得知:休息和采食是麋鹿昼间的主要活动,约占昼间时间的76%(图1)。其中:休息、采食、运动、警觉、饮水行为的时间分别占(53.40 ±5.59)%、(22.36 ±8.34)%、(11.23 ±0.63)%、(5.03 ±1.76)%、(2.65 ±0.74)%;其他行为时间占(4.42 ±2.06)%。

图1 麋鹿年均昼间活动时间分配Fig.1 Diurnal time budget activity of the Père David's deer on the yearly bass
3.2 昼间活动节律的季节变化
在不同季节,石首麋鹿昼间活动均有晨(7:00—9:00)、昏(17:00—19:00)两个采食高峰和一个明显的休息期。其中,休息期春季为10:00—15:00,冬季为11:00—15:00,持续时间相对较短;而夏季为1 0:00—1 7:00,秋季为10:00—16:00,持续时间相对较长;且在11:00—13:00和13:00—15:00两个时间段达到最高峰,期间少数麋鹿会发生警觉行为。经过早晨近2h的采食后,麋鹿会陆续走到故道河滩边及保护区低洼地形成的数个小池旁饮水,未吃饱的个体继续采食,接着再陆续地去饮水。这一行为集中在9:00—11:00时间段。接近中午时气温升高,麋鹿开始在故道河滩边、离河滩不远的草地上或树林下的阴凉处卧地休息。石首麋鹿采食频次最高的时间,春季为7:00左右(38次/min)和18:00左右(28次/min)、冬季为8:00左右(32次/min)和17:00左右(23次/min);采食频次最低的时间,夏季为7:00左右(19次/min)和19:00左右(13次/min),秋季为7:00左右(24次/min)和19:00左右(10次/min)。
3.3 昼间活动时间分配的季节变化
经Kruskal Wallis H检验,除饮水、运动在不同季节的百分比无显著差异外(P >0.05),采食、休息、警觉和其他行为在不同季节所占的百分比均有极显著差异(P <0.01)(表1,图2)。采食行为在春季和冬季的百分比相对较高;而休息行为在夏季和秋季的百分比相对较高。警觉行为和其他行为在夏季所占时间百分比最高,而在春季所占百分比最小。运动行为在秋季、冬季所占百分比相对较高。饮水行为在冬季所占百分例最小,仅为 1.83%。
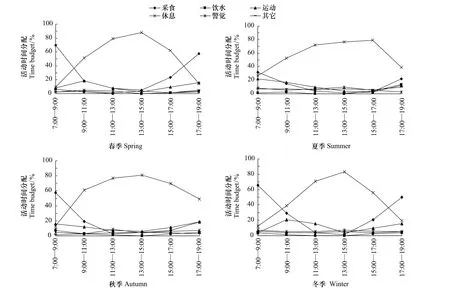
图2 不同季节中麋鹿的昼间活动节律Fig.2 Diurnal activity rhythms of the Père David's deer in different seasons
采食行为在春季和冬季占日间活动时间的百分比相对较高;而休息行为在夏季和秋季相对较高。警觉行为和其他行为在夏季的百分比最高,而在春季所占百分比最小。运动行为在秋季、冬季所占百分比相对较高。饮水行为在冬季所占百分例最小,仅为1.83%。
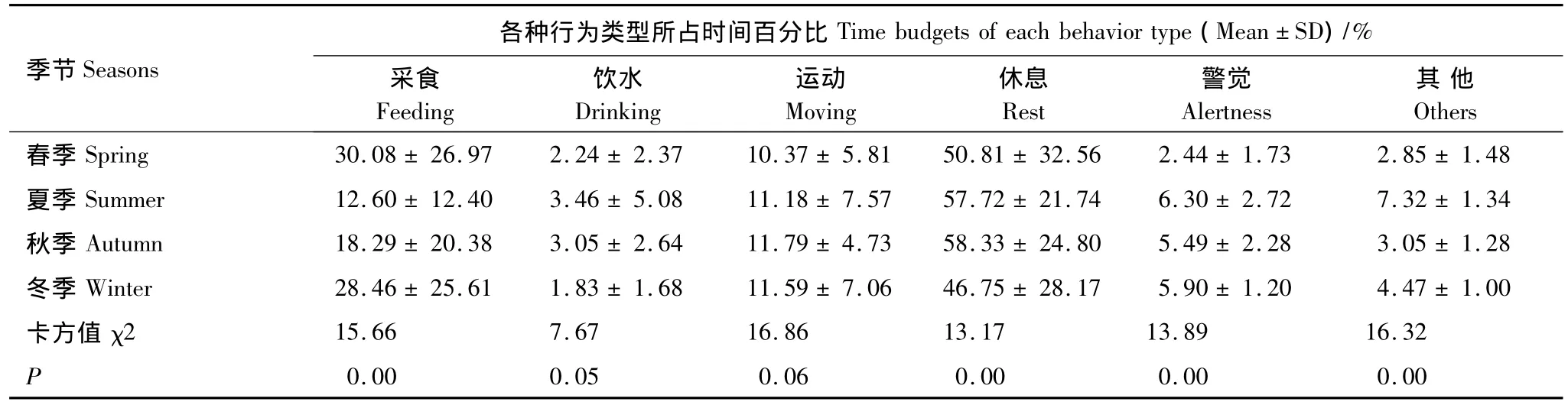
表1 麋鹿种群在不同季节中的昼间活动时间分配Table 1 Diurnal activity time budget of the Père David's deer in different seasons
3.4 不同性别-年龄组的昼间活动时间分配的差异
各性别-年龄组间运动行为有极显著差异(P <0.01),而各性别-年龄组间休息行为无显著差异(Mann-Whitney U test,P >0.05)。雄鹿与雌鹿的警觉行为有极显著差异(P <0.01),而采食、休息和其他行为无显著差异(P >0.05)。幼鹿与成年鹿及亚成年鹿的采食、运动和其他行为有极显著差异(P <0.01)。成年鹿与亚成年鹿,除运动有极显著差异外(P <0.01),其他行为无差异 (表2)。
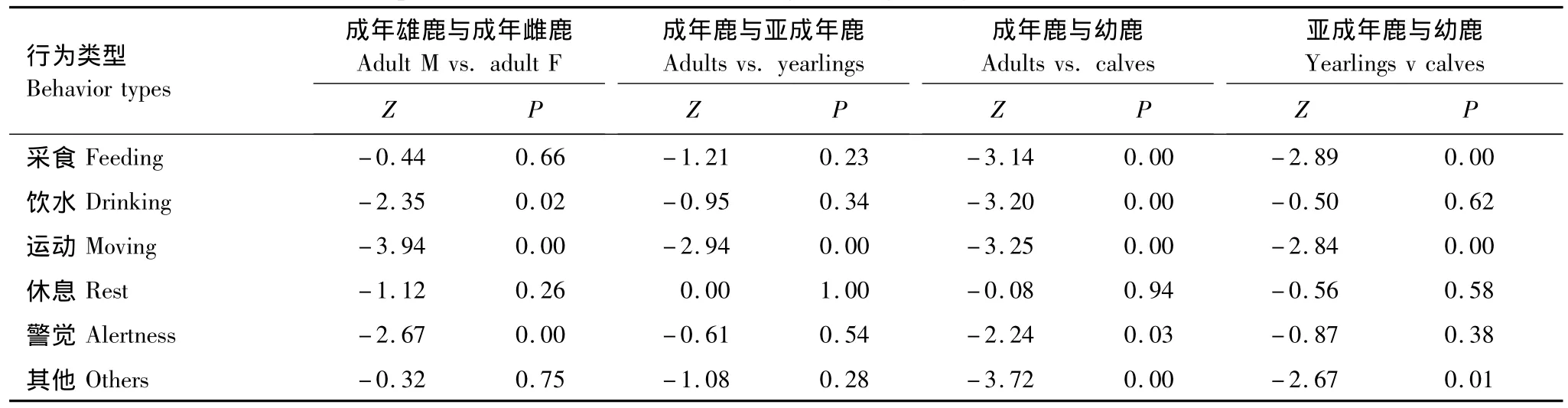
表2 麋鹿不同性别-年龄组间的各种行为时间分配比较Table 2 Comparison of diurnal activity time budget among sex-age classes of the Père David's deer
3.5 不同性别、年龄组在不同季节的昼间活动时间分配的差异
不同性别的麋鹿在不同季节的昼间活动时间有差异。无论是雄性麋鹿,还是雌性麋鹿,在春季与夏季间的采食,夏季与冬季间的饮水,春季与夏季间、夏季与冬季间运动的活动时间分配均存在显著性差异(P<0.01)(表3)。然而,雄性麋鹿夏季与春季、冬季间休息,春季与夏、秋、冬季间警觉的活动时间分配均存在显著性差异。这表明春季雄鹿用于警觉的时间最多,夏季雄鹿用于休息的时间最长。
3.6 不同季节不同年龄组昼间活动时间分配的差异
不同年龄组在不同季节的昼间活动时间分配有差异(表3)。所有年龄组,夏季与冬季饮水、春季与夏季警觉均存在显著性差异(P<0.01)。成体和亚成体麋鹿,春季与冬季警觉、夏季与冬季运动均存在显著性差异(P <0.01)。另外,成体麋鹿春季与夏季运动,亚成体麋鹿夏季与冬季休息,幼年麋鹿春季与夏季采食,春季与夏季休息,春季与夏、冬季以及夏季与秋季间饮水时间均存在显著性差异。这表明所有年龄组在夏季饮水和休息、春季警觉行为所占时间均较多;幼年麋鹿在春季比夏季采食时间明显要多。
4 讨论
4.1 昼间活动时间分配
反刍动物通常表现出“取食-休息-取食”的昼间活动规律,但取食与休息的时间并非平均分配[24-25]。反刍动物的午间休息有利于抵御太阳暴晒和反刍食物便于消化吸收。石首麋鹿昼间活动有晨、昏两个采食高峰和一个明显的午间休息期,采食和休息两种行为共占昼间活动时间的75.76%,且春季和冬季的晨、昏采食频次相对较高。这种昼间活动规律与其他反刍动物一致[26-27],梅花鹿(Cervus nippon)和岩羊 (Pseudois nayaur)等反刍动物也具有相似的昼间活动规律[28-29]。
动物的昼间活动时间分配和活动节律可间接地反映其食物的可利用性、生境质量及物种的行为学特征[29]。石首保护区麋鹿成体、亚成体、幼体全年采食行为时间分别占全年昼间活动时间的(32.12±8.36)%、(20.57 ±8.54)%、(11.13±5.28)%,这表明成年麋鹿采食需求最高。圈养麋鹿[7]与圈养马鹿(Cervus elaphus)[30]均表现出相对固定的行为模式,在集中的时间段采食,白天以休息为主,很少走动;散放麋鹿则表现出采食和运动所占活动时间较多、休息所占时间较圈养鹿群要少的特点。这些差异可能与食物组成和采食条件有关。圈养鹿群的食物以青草、饲料为主,而散放鹿群的食物组成相对复杂多样,鹿群需花更多时间寻找和处理食物,从而增加了采食时间。散放与人工圈养的海南坡鹿(Cervus eldi)之间也存在类似的差异[31]。
4.2 昼间活动时间分配的季节性变化
石首麋鹿的昼间活动时间分配表现出明显的季节性差异,通过调整活动时间分配(如减少休息时间而增加采食时间等)来满足生理需要和度过食物短缺期。麋鹿在冬、春两季的采食时间较夏、秋两季长,这与食物可利用性和气候的季节性变化有关。湿地生境中的食物呈斑块状分布,食物丰富度和可利用性随季节交替而变化;此外,降雨量和光照强度的季节性变化也明显地影响植物的生长发育[3]。春季食物充足,石首麋鹿花较多时间用于采食,以恢复漫长冬季消耗的体能,并为繁殖做好准备。冬季食物短缺,且植物的可利用性明显低于夏、秋两季,麋鹿不得不花大量的时间用于采食。据作者2004年的实地调查,保护区共有麋鹿可食植物129种,隶属35科、94属。其中,冬季麋鹿可食植物有22种,可分为二型鳞苔草(Carex dimorpholepis)、单性苔草(Carex unisexualis)、狗牙根(Cynodon dactylon)、双穗雀稗(Paspalum paspaloides)、芦苇(Phragmites communis)等冬季枯草;紫云英(Guddenstadtia multiflora)、看麦娘(Alopecurusaequalis)、水田碎米荠(Cardamine lyrate)、早熟禾(Poa pratensis)、繁缕(Stellaria media)、猪秧秧(Galium aparine)等冬绿鲜草和早春鲜草等三大类。冬季保护区内大部分植物枯萎,麋鹿可食植物种类和生物量明显减少。夏季,保护区内除小片意杨林和旱柳林可遮荫外,大部分地表均处在曝晒之下,为躲避酷暑,麋鹿大部分时间卧息于河水中或河边,很少采食;且夏季湿地植物繁盛,麋鹿只需花较少时间便可获得充足的食物。美洲马鹿(Cervus elaphus nelsoni)也采取相同的策略应对食物资源的季节性匮乏,其季节性迁徙由栖息地气候和可利用的食物量所决定[25,32]。驯鹿
(Rangifer tarandus)从春、夏季栖息地转移到冬季栖息地,也是由季节性食物丰富度所决定的[33]。
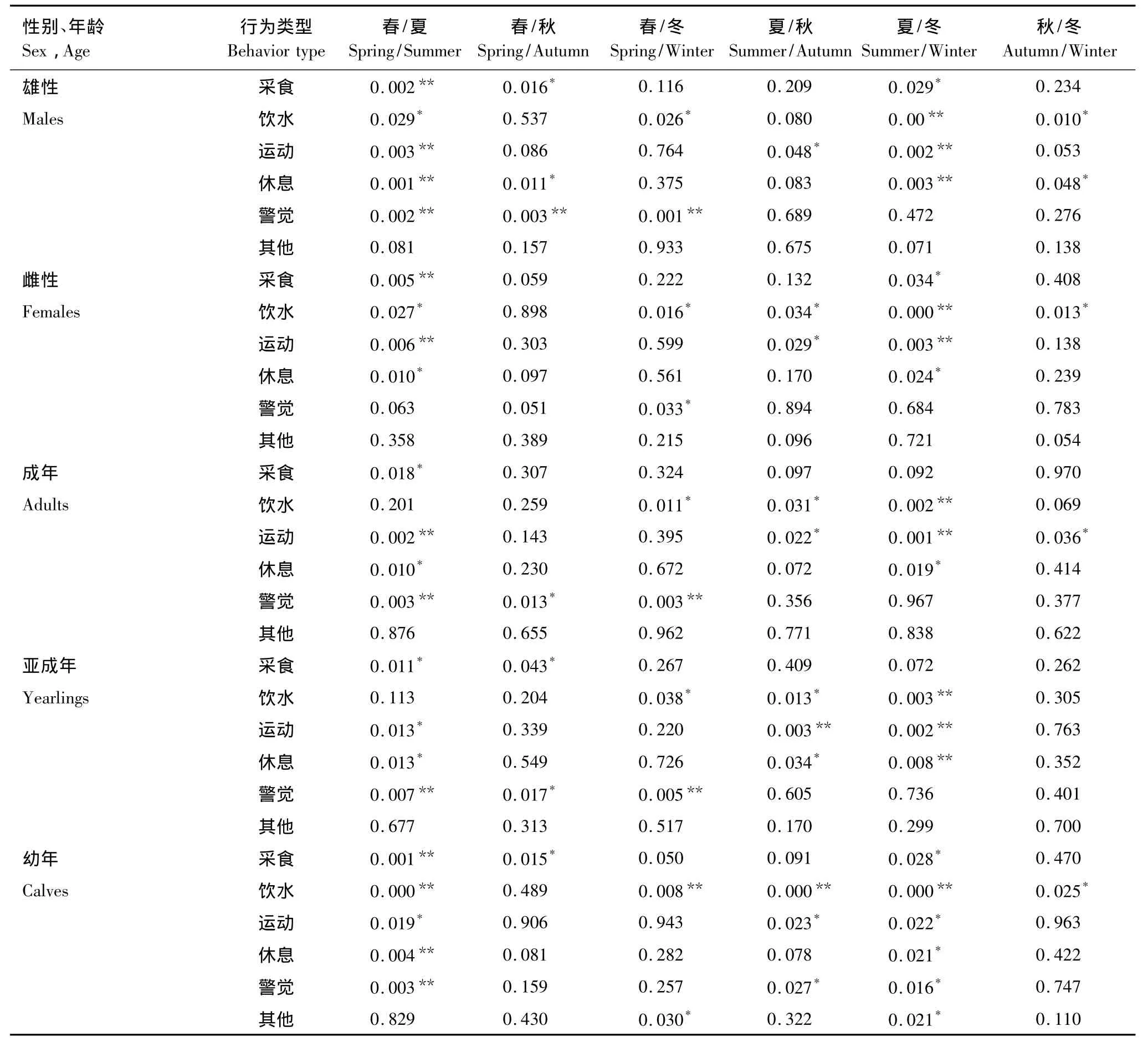
表3 不同季节不同性别年龄组的昼间活动时间分配上的差异(ANOVA分析)Table 3 Comparison of diurnal activity time budget between different sex-age classes of the Père David's deer in different seasons(ANOVA test)
石首麋鹿在夏、冬季的警觉行为所占昼间活动时间比例明显高于春季。在石首麋鹿保护区,夏季有捕鱼船在长江故道内撒网,冬季有人收割芦苇,还有人翻越进入保护区围栏捡拾鹿角,因此受人为活动的影响,麋鹿警觉行为明显地增高。雄鹿的警觉行为所占昼间活动时间比例高于雌鹿,且警觉行为主要集中在1月份、8—9月份,这与1月份有人捡拾鹿角和8—9月份的人类生产活动等干扰有关。其他行为 (排泄和繁殖)受季节因素的影响极其显著,夏季所占时间比例最高,其次是冬、秋季,春季最小。饮水和运动行为受季节和食物因素的影响不显著。
4.3 不同性别-年龄组的昼间活动时间分配
石首麋鹿在不同性别-年龄组间的运动行为差异极显著 (P <0.01)。幼鹿的运动时间多于成年鹿和亚成年鹿,这与幼鹿随时跟随着母鹿、喜嬉戏玩耍且不需担任警戒有关。幼鹿的采食行为与成年鹿及亚成年鹿差异极显著(P<0.01),这主要与它们间的采食量有明显差异有关。因成年鹿活动量大,能量消耗多,食物需求量相对较大,故采食时间长;幼鹿获取能量以母乳为主,偶尔采食,故采食时间短。幼鹿的饮水行为所占时间比最小,这主要与采食量少、喜玩耍有关。成年鹿警觉行为最多,与成年鹿担负鹿群的警戒任务有关。
因体重差别,同一物种的不同性别和年龄个体的行为时间分配也可能会存在一定差异[34]。石首麋鹿雌、雄鹿之间在休息、采食和其他行为方面的差异不显著,但在警觉、运动行为方面差异极显著(P <0.01),其原因可能是雌、雄麋鹿在繁殖策略上的差异所致。雄鹿在鹿群中一般担当警戒角色,且在繁殖期内常常为争夺与雌鹿的交配权而进行激烈争斗[3,7]。故雄鹿的警觉时间比雌鹿长。发情期雄鹿(群主和单身汉)需随时警惕其他雄鹿的挑衅,造成雄鹿的运动行为增多,运动时间较长,导致雌、雄鹿在运动时间上表现出显著的差异。这些研究结果,将对今后麋鹿迁地保护、自然野化种群恢复工作具有重要的参考价值。
致谢:中南林业科技大学野生动植物保护研究所何振、冯斌曾协助外业调查,WWF长沙办公室张琛和赵建国、湖北石首麋鹿国家级自然保护区管理处的全体工作人员给予帮助,特此致谢。
[1] Dunbar R I M.Time:a hidden constraint on the behavioral ecology of baboons.Behavioral Ecology and Sociobiology,1992,31(1):35-49.
[2] Di F A,Rodman S.Time allocation patterns of lowland woolly monkeys(Lagothrix lagotricha poeppigii)in a neotropical terra firma forest.International Journal of Primatology,2001,22(3):449-480.
[3] Jiang Z G,Li C W,Zeng Y,Widemo F.“Harem defending”or“challenging”:alternative individual mating tactics in Père David's deer under different time constraint.Acta Zoological Sinica,2004,50(5):706-713.
[4] Wemmer C,Halverson T,Rodden M,Portillo T.The reproductive biology of female Père David's deer(Elaphurus davidianus).Zoo Biology,1989,8(1):49-55.
[5] Ohtaishi N,Sheng H L.Deer of China Biology and Management.Elsever Science Publishers,B.V the Netherlands,1993:14-29.
[6] Cai G Q,Xie J H,Mathur N.Time budget and social behaviour of MI-LU(Elaphurus davidianus)in rutting season.Acta Theriologica Sinica,1988,8(3):166-171.
[7] Anwar T,Gulisi M,Patigul.The comparative study on the time budget and social behavior of David's deer.Journal of Xinjiang University(Natural Science Edition),1999,16(4):78-80,86.
[8] Ding Y H.Chinese Milu Research.Jilin Science& Technology Publishing House,Jilin,2004.
[9] Jiang Z G.Behavior coding and ethogram of the Père David's deer.Acta Theriologica Sinica,2000,20(1):1-12.
[10] Yang D D,Ma J Z,He Z,Li P F,Wen H J,Jiang ZG.Population dynamics of the Père David's deer in Shishou Milu National Nature Reserve,Hubei Province,China.Acta Zoologica Sinica,2007,53(6):947-952.
[11] Yang D D,Jiang Z G,Ma J Z,Hu H J,Li P F.Causes of endangerment or extinction of some mammals and its relevance to the reintroduction of Père David's deer in Dongting Lake drainage area.Biodiversity Science,2005,13(5):451-461.
[12] Li C W,Jiang Z G,Zeng Y,You Z Q.Rutting tactics in Père David's deer stags under different population densities and during different rut periods.Biodiversity Science,2005,13(15):424-431.
[13] Jiang Z G,Li C W,Zeng Y.Mating system,mating tactics and effective population size in Père David's deer(Elaphurus davidianus).Acta Ecologica Sinica,2006,26(7):2255-2260.
[14] Zeng Y,Jiang Z G,Li C W.Genetic variability in relocated Père David's deer(Elaphurus davidianus)populations-Implications to reintroduction program.Conservation Genetics,2007,8:1051-1059.
[15] Li C W,Jiang Z G,Zeng Y,You Z Q.A note on environmental elements as essential prerequisites for behavioral expression:A case study of Père David's deer.Applied Animal Behaviour Science,2006,102:353-359.
[16] Liu J G,XUE J H,Wang L,Ding J J,Ma W L,Liu C G,Rong Y.Habitat degradation features of Père David's Deer Natural Reserve in Dafeng of Jiangsu Province,East China.Chinese Journal of Ecology,2011,30(8):1793-1798.
[17] Li C W,Yang X B,Ding Y H,Zhang L Y,Fang H X,Tang S H,Jiang Z G.Do Père David's Deer Lose Memories of Their Ancestral Predators?PLoS ONE,2011,6(8):e23623.
[18] Li W D,Yu Q J.Extinction risk and population viability analysis of wild Père David's Deer.Pratacultural Scinence,2009,26(12):108-112.
[19] Fang S B,Zhang X S,Jia X B,An S Q,Zhou C F,Xu C.Evaluation of potential habitat with an integrated analysis of a spatial conservation strategy for David's deer,Elaphurus davidians.Environmental Monitoring and Assessment,2009,150:455-468.
[20] Martin P,Bateson P.Measuring Behavior:An Introductory Guide(Third edition).Cambridge University Press,London,2007,50-55.
[21] Lü J Q,Li B G.Diurnal activity budgets of the Sichuan snub-nosed monkey(Rhinopithecus roxellana)in the Qinling Mountains of China.Acta Theriologica Sinica,2006,21(1):26-32
[22] Alonso J A,Alonso J C.Age-related differences in time budgets and parental care in wintering common cranes.Auk,1993,110:78-88.
[23] Yao X L,Piao Z J,Li B H,Zhang J,Wang X Q,YE J,Hao Z Q.Pinus koraiensis seed consumption by rodents and birds.Chinese Journal of Applied Ecology,2008,19(8):1759-1763.
[24] Shi J B,Dunber R,Buckland D,Miller D.Daytime activity budgets of feral goats(Capra hircus)on the Isle of Rum:Influence of season,age,and sex.Cannadian Journal of Zoology,2003,81:803-815.
[25] Jiang Z G,Hudson R J.Ambient temperature,food and foraging activities of Wapiti,Cervus elaphus nelsoni,in aspen boreal forest.Acta Theriologica Sinica,1996,16(4):250-258.
[26] Clutton-Brock T H,Guinness F E,Albon S D.Red Deer:Behavior and Ecology of Two Sexes.Chicago:University of Chicago Press,1982:1-378.
[27] Green R A,Bear G D.Seasonal cycles and daily activity patterns of rocky mountain elk.Journal of Wildlife Management,1990,54:272-279.
[28] Guo Y S.Daily activity rhythm and time budget of Sichuan sika deers.Acta Theriologica Sinica,2003,23(2):104-108.
[29] Liu Z S,Wang X M,Li Z G,Cui D Y,Li X Q.Seasonal variation of diurnal activity budgets by blue sheep(Pseudois nayaur)with different Agesex classes in Helan Mountain.Zoological Research,2005,26(4):350-357.
[30] Anwar T,Arzigul K,Mahmut H.A preliminary study on the time budget and social activity pattern of captiveTarim red deer(Cervus elaphus yarkandensis).Life Science Research,2005,9(2):168-172.
[31] Wang L J,Hong M L,Chen X J.Diurnal time budget and activity rhythm of Hainan Eld's deer fawns under different breeding conditions during spring.Acta Theriologica Sinica,2005,25(1):9-13.
[32] Morgantini L E,Hudson R J.Migratory patterns of the wapiti,Cervus elaphus,in Banff National Park,Alberta.Canadian Field-Naturalist,1988,102:12-19.
[33] Baker R R.The evolutionary ecology of animal migration.London:Hodder& Stoughton,1978.
[34] Bunnell D J,Gillingham M P.Foraging behaviour:Dynamics of dining out.In:Hudson R J,White R G.Bioenergetics of Wild Herbivors.Boca Raton:CRC Press,1985:53-75.
参考文献:
[3] 蒋志刚,李春旺,曾岩,Widemo F.“占群”还是“挑战”?不同时间限制条件下麋鹿个体的交配计策.动物学报,2004,50(5):706-713.
[6] 蔡桂全,谢家华,Mathur N.麋鹿发情期主要活动的时间分配及行为研究.兽类学报,1988,8(3):166-171.
[7] 艾尼瓦尔·铁木尔,古丽司玛衣,帕提古丽.人工条件下麋鹿活动时间分配及行为的初步研究.新疆大学学报(自然科学版),1999,16(4):78-80,86.
[8] 丁玉华.中国麋鹿研究.吉林:吉林科学技术出版社,2004.
[9] 蒋志刚.麋鹿行为谱及PAE编码系统.兽类学报,2000,20(1):1-12.
[10] 杨道德,马建章,何振,李鹏飞,温华军,蒋志刚.湖北石首麋鹿国家级自然保护区麋鹿种群动态.动物学报,2007,53(6):947-952.
[11] 杨道德,蒋志刚,马建章,胡慧建,李鹏飞.洞庭湖流域麋鹿等哺乳动物濒危灭绝原因分析及其对麋鹿重引入的启示.生物多样性,2005,13(5):451-461.
[12] 李春旺,蒋志刚,曾岩,游章强.不同密度下和不同发情期时段中雄性麋鹿的繁殖计策.生物多样性,2005,13(5):424-431.
[13] 蒋志刚,李春旺,曾岩.麋鹿的配偶制度、交配计策与有效种群.生态学报,2006,26(7):2255-2260.
[16] 刘金根,薛建辉,王磊,丁晶晶,马婉丽,刘成刚,戎宇.江苏大丰麋鹿自然保护区栖息地退化特征.生态学杂志,2011,30(8):1793-1798.
[18] 李维德,于清娟.野放麋鹿种群灭绝风险与生存力分析.草业科学,2009,26(12):108-112.
[21] 吕九全,李保国.秦岭川金丝猴的昼间活动时间分配.兽类学报,2006,26(1):26-32.
[23] 姚晓琳,朴正吉,李步杭,张健,王绪高,叶吉,郝占庆.啮齿动物和鸟类对红松种子的消耗.应用生态学报,2008,19(8):1759-1763.
[25] 蒋志刚,Hudson R J.寒带针阔混交林中美洲马鹿的昼夜采食节律与食物、气温的关系.兽类学报,1996,16(4):250-258.
[28] 郭延蜀.四川梅花鹿的昼夜活动节律与时间分配.兽类学报,2003,23(2):104-108.
[29] 刘振生,王小明,李志刚,崔多英,李新庆.贺兰山岩羊不同年龄和性别昼间时间分配的季节差异.动物学研究,2005,26(4):350-357.
[30] 艾尼瓦尔·吐米尔,阿孜古丽·克依木,马合木提·哈力克.圈养塔里木马鹿白昼活动时间分配及行为格局的初步研究.生命科学研究,2005,9(2):168-172.
[31] 王力军,洪美玲,陈兴军.不同饲养条件下海南坡鹿幼体春季昼间行为时间分配及活动节律.兽类学报,2005,25(1):9-13.
