褐鳞苔草种子生物学特性研究
2013-09-08邴丹珲康建军
朱 丽,王 芳,赵 明,邴丹珲,康建军
(甘肃省林业科学研究院,甘肃兰州 730020)
褐鳞苔草(Carex brunnescens)属莎草科苔草属,是玛曲当地草地生态系统的重要建群种和优势种[1],具有适应高寒环境、生长速度快、固沙效果好、耐践踏、耐干旱等特点,对维护玛曲高寒草甸生物多样性和生态系统稳定具有重要作用[2-3]。但是,目前国内外学者对褐鳞苔草的研究报道较少,特别是对褐鳞苔草的生理生态方面的研究几乎是一片空白,加之褐鳞苔草种子具有强休眠、低萌发率等特性,致使其种子繁殖困难,自然状态下主要靠地下茎繁殖[4],从而使苗源培育困难大,不利于大面积推广并应用于固沙生产实践。因此,以褐鳞苔草种子为研究对象,通过观测其种子的形态特征、解剖特征,测定种子生活力,分析种子形态、生活力与种子萌发之间的关系,了解其生物学特性,不仅能更深刻地认识褐鳞苔草,而且对玛曲高寒牧区植被生态恢复具有重要意义。
1 试验材料及方法
1.1 试验材料
试验所用的褐鳞苔草种子于2011年7月采自玛曲县试验点,该试验点位于玛曲黄河大桥以西,地处玛曲县欧拉乡欧强村,距县城8.5 km。
1.2 试验方法
1.2.1 种子的形态特征
通过对形状、颜色、大小和质量的观测,描述褐鳞苔草种子的基本特征。形状和颜色采用观察法描述;大小采用0—150 mm电子数显卡尺测定长轴、短轴和厚度;质量参照中华人民共和国国家标准《牧草种子检验规程》(GB/T 2930.5—2001),采用百粒法(即从纯净种子中随机抽取100粒种子为1组,重复取8组称量)测定种子千粒重。
1.2.2 种子处理方法
由于褐鳞苔草种子种皮坚硬,很难切割,不做任何处理情况下种子几乎不吸水,因而首先要对种子进行处理(见表1),然后解剖观察、测定生活力。

表1 褐鳞苔草种子处理方法
1.2.3 种子的解剖特征
待处理过的种子种皮变软后随机选取30粒种子进行解剖,在立体显微镜下观察胚的发育情况。
1.2.4 种子的生活力测定
采用TTC染色法测定种子的生活力。对处理过的种子采用整粒染色和纵切胚之后再染色的方法,用0.5%的四唑溶液在35℃、黑暗条件下染色24 h,然后清水冲洗,在立体解剖镜下观察,根据染色的部位和染色程度,参照《牧草种子检验规程》(GB/T 2930.5—2001)鉴定种子有无生活力。
2 结果与分析
2.1 不同处理对种子的影响
所有处理中,只有经浓H2SO4和NaOH浸泡过的种子浸出液颜色较深,种皮、种子软化,容易切割,其余各处理无变化,种皮坚硬,很难切割,种子几乎没吸水。种子萌发情况见表2。
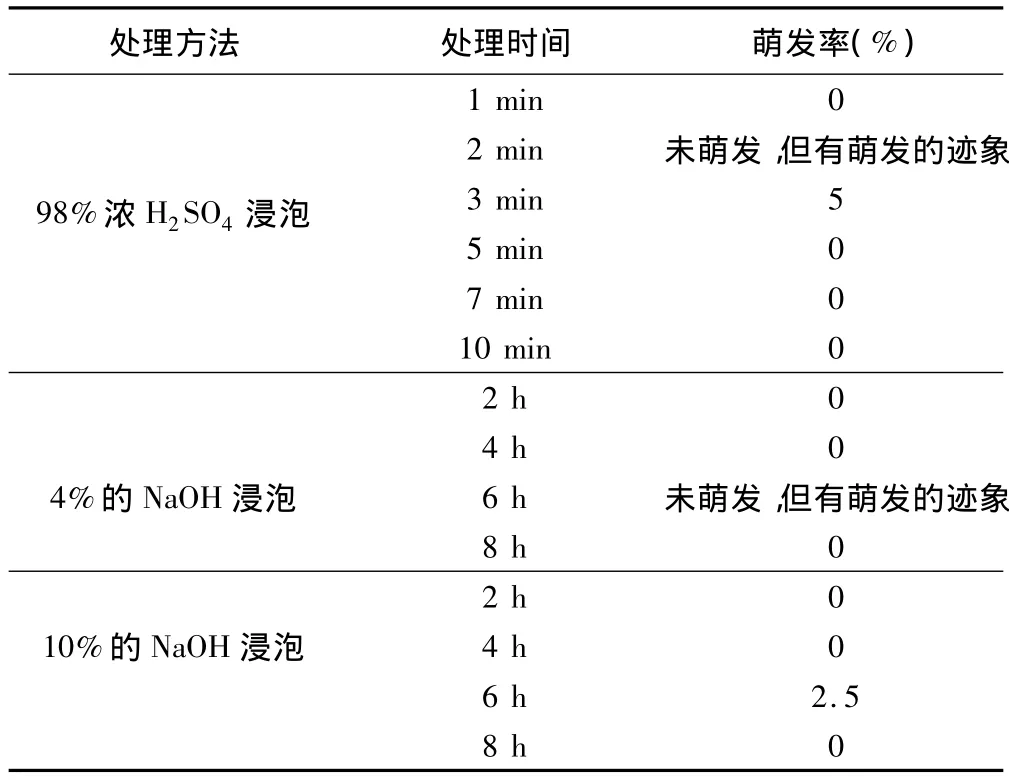
表2 浓H2SO4和NaOH对褐鳞苔草种子萌发率的影响
随着处理时间的增加,经浓H2SO4处理的褐鳞苔草种子浸出液的颜色由浅变深,当处理时间达到4 min时,颜色已经变成黑色,处理10 min时,种子已经完全炭化,失去生命活力。研究结果表明,处理3 min时种子萌发能力较其他处理有所提高。
由此可见,用浓硫酸处理种子,在一定时间内发芽率与处理时间呈正相关。这是由于浓硫酸对种皮的腐蚀需要一定的时间,其时间长短与种皮的结构有关。当酸腐蚀种皮后,就可解除种皮障碍,打破休眠,提高发芽率;而当处理时间延长至整个种皮崩溃、伤及胚时,发芽率下降;随着时间的增加,胚的受损伤程度加大,此时发芽率与处理时间呈负相关。因此,浓H2SO4的处理时间应使种皮结构受到破坏而又未伤及胚为最好,此时可增加种皮通透性,有利于种子吸水膨胀而提高其发芽率[5]。
用4%的NaOH处理6 h种子有萌发的迹象,但未萌发;用10%的NaOH处理6 h的种子萌发了,但是萌发率极低,仅为2.5%;随着NaOH浸种时间延长,种子浸出液颜色逐渐加深,8 h时种子已失去发芽能力。
2.2 褐鳞苔草种子的形态特征
褐鳞苔草种子呈褐色,三棱形,具有或长或短的喙,种皮硬。种子较小,长度2.32~2.46 mm,宽度1.35~1.43 mm,厚度1.23~1.25 mm,千粒重1.731 1 g。
2.3 褐鳞苔草种子的解剖特征
成熟的种子由种皮、胚和胚乳三部分组成。通过对30粒褐鳞苔草种子的解剖观察,发现有20粒种子具有完全的胚结构,即66.67%的种子胚发育完全,其余10粒为空秕种子。虽然种子已经达到成熟种子的大小、颜色,但经过筛选的种子仍有空秕,这可能是种子形成过程中受到不利环境条件的影响,使种子的内部物质积累受阻,不能继续正常发育造成的,也有可能是因为褐鳞苔草种皮厚而且较硬,采用蒸馏水漂浮法无法将空秕粒完全除去。
2.4 褐鳞苔草种子的生活力测定
经TTC染色法测定,30粒褐鳞苔草种子中有17粒的种胚大部分被染成红色,6粒染色非常浅,其余7粒根本不染色。而染色较浅或根本不染色的种子被认为属于不能正常发育的种子。由此可见,褐鳞苔草种子的生活力较低,平均为56.67%,这说明玛曲高寒草甸的褐鳞苔草种子一半以上具有萌发的潜力。
3 讨论
3.1 褐鳞苔草种子的形态特征是长期适应生态环境的结果
种子休眠是植物为了适应环境而在长期演化过程中获得的一种生物学适应,它对“种”的保存、繁衍是极为有利的。因此,通常情况下除顽拗性种子外[6],大多数植物种子或强或弱地具有休眠特性,尤其以寒带和温带的植物种类更为明显。研究表明,褐鳞苔草种子小而且种皮坚硬,这是因为褐鳞苔草产于中国—喜马拉雅—中亚高山的高寒草甸,长期生长在仅有冷暖二季之别、温度年较差小而日较差悬殊、太阳辐射强烈的高海拔地区,形成了一套长期适应高寒生境的形态特征和生态特征[7]。这与魏疆等[8]学者的研究结论“植物的生长规律受自身生物学特性和生境条件的制约”一致。至于海拔对种子萌发的影响,目前为止还没有得出一个关于种子萌发能力与海拔之间关系的统一结论[9]。Neuffer[10]研究发现采自阿尔卑斯山不同海拔的荠菜(Capsella bursapastoris)种子的萌发与采集生境之间没有什么关系;Vera[11]在研究海拔对西班牙北部的几种石楠种子萌发的影响效应时发现,采自较高海拔的种子具有较高的萌发率;而 Miller和 Cummins[12]在研究苏格兰一种石楠(Calluna vulgaris)种子萌发时发现,采自低海拔的种子具有较高的萌发率。
3.2 褐鳞苔草种皮表面结构有可能是阻碍种子萌发的障碍之一
褐鳞苔草的坚硬种皮透水性和透气性差,使胚生长得不到充足的水分和氧气,从而抑制了种子的萌发。李曜东等研究发现埋存于地下的古莲(Nelumbo nucifera)种子休眠可达几百年甚至上千年,坚硬的种壳可阻止氧气进入是重要原因[13-14]。种皮的存在对胚的发育常常起到机械的阻碍作用,反枝苋(Amaranthus retroflexus)等植物种皮虽然对水分有透过性,但种皮非常坚固,使胚根不能穿破种皮,从而抑制萌发[15]。藏嵩草(Kobresia tibetica)完整种子室内和野外萌发率分别为4%和2%,而剥去种皮后种子萌发率达47%[16],种皮坚硬是造成种子萌发率低的主要原因。
种皮附属物以及生长抑制物质对种子萌发与休眠也是有影响的。有些荒漠植物果皮或种皮外包有黏液层,这种结构对种子的萌发及幼苗生长有促进作用[17],如白沙蒿(Artemisia sphaerocephala)成熟瘦果上的黏液物[18];但是有些植物的种胚覆盖物含有生长抑制物质而抑制种子萌发,例如褐鳞苔草。解剖镜下观察发现,褐鳞苔草的种皮表面光亮、质地坚实、结构致密,并且嵌有数量较多的类晶体物质,褐鳞苔草种皮表面的这种质地与结构,可能会对萌动种子产生较大的机械束缚力,进而限制到褐鳞苔草种子的萌发。
4 结论
褐鳞苔草为了适应气候恶劣的高寒草甸环境,弱化了种子的繁殖能力,自然状态下几乎不萌发。褐鳞苔草在长期适应生态环境过程中形成种子生活力较低、种子小且种皮坚硬的形态特征以及生理特征抑制种子萌发。经浓H2SO4和NaOH处理虽能破坏种皮结构,促使种子吸水、透气,但是种子萌发率极低,仍不利于大面积推广使用。因而,对于具有萌发潜力的褐鳞苔草种子萌发所需的适宜条件还有待进一步深入研究。
[1]张怀山,杜文华,田福平,等.甘南高原沼泽植物群落类型与组成特征研究[J].中国草地学报,2008,30(4):64-67.
[2]魏强,王芳,陈文业.黄河上游玛曲不同退化程度高寒草地土壤物理特性研究[J].水土保持通报,2010,30(5):16-21.
[3]王一博,王根绪,沈永平.青藏高原高寒区草地生态环境系统退化研究[J].冰川冻土,2005,27(5):633-640.
[4]吉文丽,朱清科,李卫忠,等.苔草植物分类、利用及物质循环研究进展[J].草业科学,2006,23(2):15-21.
[5]葛俊彦,蔡亚非.不同处理方法对几种牧草种子发芽率的影响[J].资源开发与市场,2009,25(1):4-6.
[6]杨期和,尹小娟,叶万辉,等.顽拗型种子的生物学特性及种子顽拗性的进化[J].生态学杂志,2006,25(1):79-86.
[7]张怀山,张吉宇,乔国华,等.黄河首曲——玛曲湿地沼生植物的群系分类研究[J].湖北农业科学,2011,50(5):924-926.
[8]魏疆,张希明,单立山,等.梭梭幼苗生长动态及其对沙漠腹地生境条件的适应策略[J].中国科学D辑:地球科学,2006,36(z2):95-102.
[9]卜海燕.青藏高原东部高寒草甸植物种子的萌发与休眠研究[D].兰州:兰州大学,2007:9-10.
[10]Neuffer B,Bartelheim S.Gen-ecology of capsella bursa-pastoris from an altitudinal transsect in the Alps[J].Oecologia,1989,81(4):521-527.
[11]Vera M L.Effects of altitude and seed size on germination and seedling survival of heathland plants in north Spain[J].Plant Ecology,1997,133(1):101-106.
[12]Miller G R,Cummins R P.Role of buried viable seeds in the recolonization of disturbed ground by heather(Calluna vulgaris[L.]Hull)in the Cairngorm mountains,Scotland,UK[J].Arctic and Alpine Research,1987,19(4):396-401.
[13]李曜东,魏玉凝,徐本美.古莲子与现代莲子ABA含量和SOD活性的比较研究[J].植物学通报,2000,17(5):439-442.
[14]秦淑英,唐秀光,王文全,等.药用植物种子处理研究概况[J].种子,2001(2):37-39.
[15]曹仪植,宋占午.植物生理学[M].兰州:兰州大学出版社,1998:181-183.
[16]邓自发,谢晓玲,王启基,等.高寒草甸藏嵩草种群繁殖对策的研究[J].应用与环境生物学报,2001,7(4):332-334.
[17]马骥,李俊祯,孔红.我国沙区6种蒿属植物的种子微形态特征[J].中国沙漠,2002,22(6):586-590.
[18]黄振英,Gutterman Yitzchak,胡正海,等.白沙蒿种子萌发特性的研究Ⅰ.粘液瘦果的结构和功能[J].植物生态学报,2001,25(1):22-28.
