有机锡致海洋腹足类性畸变分子机制的研究进展
2013-08-20肖丽萍王淑红邹志华王艺磊张子平
肖丽萍,王淑红,*,邹志华,王艺磊,张子平
1.集美大学水产学院,厦门361021
2.西东大学生物系,美国新泽西州07079
有机锡如三丁基锡(tributyltin,TBT),作为防污损生物附着的添加剂自20世纪60年代就广泛应用于轮船、码头等涂料中,但在防污的同时对非靶生物也具有短期和长期的毒性效应,如可对哺乳动物的生殖系统、免疫系统以及神经系统等造成不同程度的损伤[1],也可导致草履虫[2]、大型蚤[3]、腹足类[4]等水生无脊椎动物的性畸变和死亡。其中,最显著的生态毒理学效应是诱发海洋腹足类的性畸变(imposex),由此导致的腹足类种群的衰退是目前海洋污染事件中最沉重的教训[5]。性畸变是指软体动物雌性个体产生不正常的雄性特征,如阴茎和输精管的形成;性畸变严重时,会导致雌性个体生殖能力的丧失,造成种群衰退甚至局域性灭绝[6]。性畸变的研究最早起源于1970年在狗岩螺(Nucella lapillus)的报道,目前估计全世界发生性畸变的种类至少在200种以上,其中新腹足类有50多种[7-11]。
有机锡污染是一种典型的海洋污染事件,常规的物理和化学等监测方法都存在操作复杂、不易观察和辨别、灵敏性低和成本高等局限性,因而很多学者建议采用某些腹足类性畸变状况来监测不同海域有机锡的污染现状。Gibbs等[12]根据狗岩螺畸变个体阴茎、输精管和雌性生殖道的损伤程度将其划分为6个不同的发展阶段,并提出阴茎相对大小指数(relative penis size index,RPSI)、输精管发展指数(vas deferens sequences index,VDSI)量化的方法来评估种群受影响的程度。之后,Shi等[7]根据畸变物种差异性提出了普适的性畸变划分图来更准确地反映性畸变的形态变化及程度评估。一些学者应用这些形态学指标相继调查了世界各个海域腹足类的性畸变状况[13-18]。利用腹足类性畸变的形态学变化监测评估有机锡的污染是目前海洋环境生态学监测的最成功范例,腹足类性畸变已成为内分泌干扰物在种群水平影响生态平衡的重要证据[19-20],对类似内分泌干扰物的风险评价和管理意义重大。因此,腹足类的性畸变可作为有机锡生物监测的有效指标[4],并成为促使2003年国际海事组织全面禁止船体上涂含有TBT的防污漆的禁令颁布的主要原因和评判法规效果的重要标尺。
尽管腹足类性畸变形态学指标在有机锡污染监测中发挥了重要的作用,但该类指标的应用仍存在一些问题。首先,性畸变形态学特征存在种属差异,且这种方法仅适用于中腹足目和新腹足目的种类,对无明显性别特征的原始腹足类如鲍等则不适用;最重要的是,形态学监测手段只能检测到较晚期的性畸变(往往已不可逆转),无法预警预报,也难以达到保护生物种群的目的。因此,有必要进一步深入研究有机锡致腹足类性畸变的作用机制,进而筛选敏感的早期检测指标。虽然腹足类性畸变的形态学变化已被广泛用于监测有机锡的污染状况,但其分子机制一直众说纷纭,主要原因在于软体动物的性别决定机制至今尚未明确,性别决定和性别分化相关基因和蛋白尚未得以证实[21-24],相关研究的滞后大大增加了腹足类性畸变分子机制研究的难度。然而近年来,无论是传统的脊椎动物类型的类固醇激素假说,还是较新的视黄酸X受体(retinoid X receptor,RXR)假说,都有了新的进展,尤其是 RXR受体假说,涌现了大量有价值的文献。论文将结合近期的研究进展,对有机锡诱发腹足类性畸变分子机制的研究进行综述,以便更全面地了解其致毒机理,为有机锡及其他内分泌干扰物的早期快速监测和重要水产经济资源的保护提供理论依据。
1 脊椎动物类型的类固醇激素假说
该假说认为腹足类体内存在着类似于脊椎动物类型的性激素——睾酮和雌二醇,且腹足类的性别调控也类似于脊椎动物,由上述2种性激素调控。有机锡可通过影响性激素的合成或代谢过程而改变其在体内的正常水平,进而导致性畸变。相继有学者采用气相色谱-质谱联用技术[25-26]和放射免疫分析技术[27-28]证实并测定了这2种激素在腹足类和其他软体动物中的存在。有关东泥织纹螺(Ilyanassa obsoleta)[29]、大洋角螺(Marisa cornuarietis)[30-31]、狗岩螺(Nucella lapillus)[32-34]、新西兰泥螺(Potamopyrgus antipodarum)[35]的一些研究也似乎表明,这些激素在腹足类生殖和性畸变诱导中发挥着作用,但具体机制尚未阐明,相关的研究主要涉及性激素合成的关键酶P450芳香化酶和影响游离睾酮浓度的一些因素。
1.1 芳香化酶抑制假说
P450芳香化酶是脊椎动物负责将雄激素芳香化为雌激素的关键酶,该假说认为,TBT能抑制P450芳香化酶的活性,造成睾酮在体内的积累而引起性畸变[32]。邓瑞鹏等[36]通过细胞色素 P450,b5和细胞色素P450还原酶的含量测定发现,性畸变疣荔枝螺(Thais clavigera)P450酶系的这3种组分与正常雌性螺有显著差异,而与雄性螺更为接近;同时,Santos等[27]等进一步将狗岩螺暴露于芳香化酶抑制剂4-羟基-睾酮下,间接证明TBT可竞争性地抑制腹足类体内P450芳香化酶的活性而诱发性畸变,但不能增加其严重程度。然而迄今为止,并无直接证据表明芳香化酶在无脊椎动物类固醇激素代谢过程中的作用;并且在对生蛤仔(Ruditapes decussatus)[37]和欧洲玉黍螺(Littorina littorea)[38]的研究中发现,芳香化酶在睾酮代谢过程中只占极小的比例。更值得指出的是,脊椎动物类型的P450芳香化酶基因CYP19至今尚未在脊索动物门之外的无脊椎动物中被证实[21]。因此,很难用芳香化酶抑制假说来解释睾酮在腹足类一些种类体内的积累。
1.2 游离睾酮水平调节假说
该假说认为,TBT能够影响睾酮与脂肪酸的结合,通过影响游离睾酮的浓度而导致性畸变。Gooding 和 LeBlanc[39]以及 Janer等[28,40]的研究表明,在某些软体动物,睾酮和雌二醇与脂肪酸的酯化结合是调控这些性激素水平的主要代谢途径。Gooding等[41]的研究表明,东泥织纹螺能够以脂肪酸酯化物的形式生物转化和储存多余的睾酮,并发现不同TBT浓度下东泥织纹螺体内总睾酮含量不变而游离睾酮含量有所上升。Abidli等[42]研究了2种TBT敏感物种环带骨螺(Hexaplex trunculus)和染料骨螺(Bolinus brandaris)在TBT暴露下类固醇激素(雌二醇和睾酮)的含量变化,进一步证明这2种性激素都同时以游离和酯化的形式存在于消化腺和性腺复合体中,且TBT暴露会引起雌性染料骨螺游离睾酮水平增加而酯化睾酮减少,而在雌性环带骨螺中游离和酯化的睾酮水平都会显著地上升,但TBT对这2种螺的雄性个体的睾酮和雌二醇水平都没有影响。该研究表明TBT暴露会引起雌性个体游离睾酮增加,导致性激素比例失衡而诱发性畸变,并且TBT在不同物种、不同性别中对性激素代谢的影响存在差异。
尽管上述研究在腹足类组织中能检测到类似于脊椎动物的雌雄激素,且有机锡的暴露干扰了某些种类体内类固醇激素的水平,但由于并无直接证据表明软体动物的性别调控机制与脊椎动物类似,且NCBI核苷酸数据库中至今尚无脊索动物门之外的无脊椎动物CYP19基因的数据,也尚无学术文章报道在无脊椎动物中分离获得芳香化酶蛋白[22]。此外,在性激素受体研究方面,尽管类雌激素受体cDNA 已在疣荔枝螺[43]和东泥织纹螺[44]中被分离出来,但其编码的蛋白是一种组成型转录激活因子,且雌激素并不能与该蛋白结合;同时,尚无任何有关成功克隆包括腹足类在内的无脊椎动物雄激素受体的研究报道。Scott[23-24]在最新的综述中指出,在软体动物体内检测到类似于脊椎动物性激素的现有报道不能排除这些激素来源于其生活的水体环境而非软体动物自身合成的可能性。因此,尽管腹足类体内存在类似于脊椎动物的雌激素和雄激素,但由于缺乏对其生物合成途径或来源的了解,以及有关受体的直接证据,由此引出的脊椎动物类型的类固醇激素假说难以获得有力支持。
2 神经肽假说
该假说认为,TBT能经由退化因子作用于脑神经节而导致神经肽阴茎形态发生因子(penis morphogenetic factor,PMF)的异常释放,其中PMF负责雌性个体中阴茎的形成[45]。Féral和 LeGall[46]通过组织培养方法发现,TBT暴露后提取得到的西欧骨螺(Ocenebra erinacea)神经中枢能诱发未成熟的舟螺(Crepidula fornicata)阴茎形成区发育成为阴茎,且与神经系统的发育同步,表明TBT能作为神经毒素改变神经激素如PMF的分泌而引起性别变异。目前有学者已在狗岩螺(Nucella lapillus)[47]、盘大鲍(Haliotis gigantea)[48]等个体的中枢神经系统中发现TBT和三苯基锡(triphenyltin,TPT)的累积,表明TBT和TPT可能对神经内分泌系统发挥毒性效应。此外,Oberdörster和 McClellan-Green[29]报道了腹足类中枢神经节分泌的一种神经肽APGWamide(Ala-Pro-GLy-Trp-NH2),作为一种假定 PMF,能显著诱发雌性东泥织纹螺性畸变;之后该作者进一步证实了APGWamide是腹足类东泥织纹螺的一种PMF,并且该物质在性畸变个体和雄性个体中分布模式相似,都主要聚集于内脏团[45]。然而,在上述研究中,由于TBT或TPT暴露导致过高的性畸变率和超长的阴茎,有可能弱化了APGWamide诱导或促进性畸变的作用;Santos[49]和 Castro 等[50]的研究却发现,APGWamide不能诱导染料骨螺和狗岩螺性畸变,而目前TBT暴露、神经激素如神经肽APGWamide异常释放和性畸变之间的因果关系尚未有直接证据来证实,所以该假说也有待进一步的深入研究。
3 RXR假说
RXR是配体激活的转录因子核受体超家族成员,有α、β、γ共3种亚型,每种亚型分别由不同的基因编码,可作为其他核受体异二聚体伙伴或同源二聚体在核受体调控信号通路中发挥作用,参与细胞生长、分化、代谢和胚胎发育等调节[51]。近年来,越来越多的证据表明,RXR参与TBT诱导腹足类性畸变过程。
Nishikawa 等[52]通过注射 9-顺式维甲酸(9-cis retinoic acid,9cRA)、TBT和TPT,发现注射9cRA 会诱发雌性疣荔枝螺性畸变,在性畸变严重个体中RXR表达显著增加,并利用酵母双杂交系统检测到有机锡能增强hRXRs与9cRA的作用。之后,Bouton等[53]克隆得到了光滑双脐螺(Biomphalaria glabrata)RXR同系物;Castro等[50]克隆了狗岩螺RXRa和RXRb两种亚型,并再次证实TBT与9cRA都能诱发性畸变。Horiguchi等[54]采用实时定量PCR、免疫印迹和免疫组织化学方法,发现雄性及发生性畸变的雌性疣荔枝螺阴茎中的RXR基因表达明显高于正常雌性个体,表明RXR可能参与雌性腹足类在有机锡暴露下诱导雄性生殖器官(阴茎和输精管)生成的机制;随后该作者通过检测TPT暴露下疣荔枝螺雌雄个体各组织中RXR表达水平,发现雌性个体阴茎的增长与阴茎形成区RXR的表达增强有关联,而在雄性个体中RXR表达没有显著变化,但其最高表达量也位于阴茎处,表明RXR对腹足类雄性及性畸变雌性个体的雄性生殖器官的发育起着重要作用[55]。Urushitani等[56]克隆和鉴定了疣荔枝螺RXRs两种亚型,发现其氨基酸序列与哺乳动物和腹足纲其他种类的RXRs具有高度相似性,同时经过磷酸化位点突变检测发现9cRA应激下RXR亚型的转录活性显著地高于对照组,并经由顺式重复序列1(direct repeat 1-type,DR1)反应元件使得转录活性发生变化,表明维甲酸可能在腹足类生殖器官发育及成分组成中发挥重要作用,并且亚型间差异可能是软体动物内分泌系统调控的功能基础。此外,张纪亮等[57]综述有机锡能够作为RXR与过氧化物酶体增殖物激活受体(peroxisome proliferatoractivated receptor,PPAR)的激动剂引起螺类性畸变、促进脂肪分化并影响脂质代谢平衡。
Lima等[58]认为,TBT暴露会使不同组织、不同性别中RXR转录情况存在差异而使腹足类雌性在畸变高级阶段中RXR转录模式与雄性相同而产生雄性特征,并提出TBT诱发性畸变机制的RXR信号通路假说。该假说认为,正常阴茎分化过程中,类视黄素衍生物结合RXR后通过与RXR反应元件相互作用来调节雄性中枢神经系统(the central nervous system,CNS)的靶基因,PMF编码基因受RXR信号的控制并在雄性阴茎分化过程中被诱导,而PMF的释放会启动级联事件从而形成阴茎和输精管。在正常雌性中,PMF编码基因在缺少RXR配体的情况下会被抑制而阻断释放;而TBT会异常结合雌性CNS中的RXR从而激活PMF编码基因的信号通路,进而在雌性体内诱发雄性阴茎分化的一系列级联事件,最终形成阴茎,导致性畸变。性畸变起始可能独立于性腺和/或消化腺,但由于TBT结合RXR可能会影响性腺、消化腺中类视黄素和类固醇代谢组件的编码基因,导致这些激素水平失衡,转而调控雌雄个体CNS和雄性生殖器的RXR信号通路。
近几年,随着性畸变相关研究的不断深入,RXR假说逐渐成为性畸变机制研究的热点。Sternberg等[10]综述了腹足类性别分化及生殖调控的机制,认为一些腹足类性腺的恢复即生殖特征和第二附属器官的再生长主要受季节调控,相关的环境因素通过刺激神经及内分泌系统的一系列反应而调控雌雄个体的性别分化、产卵、精子发生及交配行为。调控性腺恢复的主导环境因素是光照周期,其次是控制繁殖和产卵时间的温度、营养状况和寄生虫等因素。TBT诱发性畸变,即阻断雌性个体性腺及性别特征的显性性别决定,这种因环境因素诱发或修改生殖过程的现象,会使腹足类不断出现生理、行为和形态方面的适应性变化,以保证后代生存几率的最大化。Gooding等[59]的研究也观察到TBT暴露只对处于性腺恢复的临时窗口期的东泥织纹螺产生作用,会刺激雌性个体的雄性特征发育,这从另一层面说明了TBT诱发腹足类性畸变的特殊性。上述研究表明,腹足类的性别分化很可能存在着关键的窗口期,光照、温度、营养状况以及某些特殊的环境内分泌干扰物如有机锡的存在,有可能彻底改变生物原来的性腺发育途径,出现性畸变或性逆转,对传统生殖系统发育理论提出了挑战。
图1综合了本文论及的与有机锡致海洋腹足类性畸变分子机制相关的a、b、c三类假说。a为脊椎动物类型的类固醇激素假说,可分为芳香化酶抑制假说和游离睾酮水平调节假说。P450芳香化酶抑制假说认为TBT能抑制该酶的活性,阻断雄激素芳香化为雌激素,造成雄激素在体内积累而引起性畸变。游离睾酮水平调节假说认为TBT能抑制乙酰辅酶A与乙酰转移酶进而阻碍睾酮与游离脂肪酸结合,引起雌性个体游离睾酮增加,导致性激素比例失衡而诱发性畸变。b为神经肽假说,该假说认为TBT能经由退化因子作用于CNS而导致PMF如神经肽(APGWamide)的异常释放,致使阴茎发育而产生性畸变。c为RXR假说,该假说认为TBT会结合雌性CNS中的RXR,通过与RXRRE(视黄酸X受体反应元件)相互作用而激活PMF的信号通路,进而诱发阴茎分化而导致性畸变;同时,RXR的激活与软体动物性腺的发育状态密切相关,腹足类如果在关键窗口期暴露于TBT,则诱导性畸变。
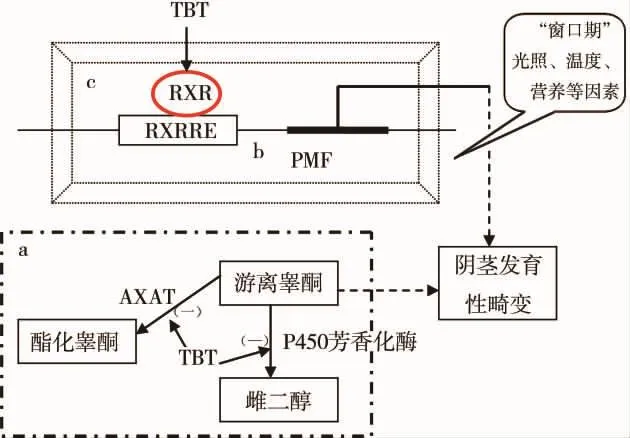
图1 有机锡致海洋腹足类性畸变的分子机制Fig.1 Molecularmechanism of organotin-induced imposex inmarine gastropods
4 腹足类性畸变个体抗氧化防御系统的变化
李张伟等[60]检测了TBT暴露下疣荔枝螺性畸变个体超氧化物歧化酶(superoxide dismutase,SOD)、脂质过氧化物(lipid peroxide,LPO)及过氧化氢酶(catalase,CAT)活性与正常雌雄个体的差异,结果表明,这几种生化指标的差异随着性畸变严重程度的加深而愈加明显,并呈现一定的规律性,其中SOD活性在正常雌性和性畸变初期阶段个体中与雄性的接近,在性畸变严重个体中则显著下降,而CAT活性在性畸变初期有上升趋势,在性畸变严重个体中则显著下降,这表明,腹足类体内重要酶在有机锡污染下会出现明显紊乱。随后该作者又研究了疣荔枝螺雄性、雌性及性畸变个体在不同发育阶段的总蛋白和酯酶同工酶水平,结果表明,性畸变个体体内蛋白表达量和酯酶活性与雄螺的相近[61]。
5 腹足类性畸变个体DNA的变化
研究表明,有机锡还具有遗传毒性,主要表现为基因突变[62]、染色体畸变[63]和 DNA 损伤[64-65]等。管云雁[66]通过对疣荔枝螺和黄口荔枝螺(Thais luteostoma)核型分析,发现性畸变个体的染色体形态有所改变而数目没有变化,并推测这种变化可能与性畸变现象有一定的相关性。同时,周小朋等[67]采用随机扩增多态性DNA技术来研究疣荔枝螺性畸变机理,结果表明,差异性片段的出现与有机锡的污染具有一定的相关性,认为有机锡的存在已引起疣荔枝螺遗传物质DNA的变化,并且这种变化是随机的。Hagger等[68]应用线性回归分析揭示了性畸变和DNA损伤程度具有紧密的相关性,同时通过组织学方法检测到狗岩螺性畸变个体输精管和阴茎的增生性生长,表明环境暴露、遗传响应和生殖变异之间存在着复杂的相互作用。
6 基因组学尺度对性畸变机制的研究
随着国内外有关腹足类性畸变的研究日渐增多和分子生物学技术的迅速发展,一些学者采用基因组学方法来进一步研究有机锡的致毒机制。Iguchi等[69]应用生态毒理基因组学方法研究脊椎及无脊椎动物不同物种如老鼠、美洲钝吻鳄、两栖动物、鱼类、腹足类和大型蚤等个体的雌激素响应基因及其表达变化,探讨了内分泌干扰物致毒的分子机制。贾锡伟等[70]构建了杂色鲍(Haliotis diversicolor)在TBT暴露诱导下的均一化cDNA文库,得到3 048个高质量表达序列标签,之后作者采用实时荧光定量PCR方法进行了相关功能基因的表达分析并得到纤维素酶、β-1,4-内葡聚糖、铁蛋白亚基1等几种较好的 TBT 监测候选基因[71]。Pascoal等[72]对 TBT暴露狗岩螺的转录组测序数据分析再次支持了上述3类假说所涉及的性激素、神经多肽和RXR参与有机锡诱发腹足类性畸变过程,同时通过PPARγ竞争抑制剂罗格列酮的体内注射实验,证实无脊椎动物包括腹足类的内分泌干扰过程中存在着与脊椎动物共通的 PPAR 信号通路。Titley-O'Neal等[73]采用基因芯片技术比较了大不列颠维京岛附近不同站位女皇凤凰螺(Strombus gigas)的基因表达谱,发现了高浓度TBT暴露站位差异表达的17个基因,功能分析表明,钙离子结合、免疫反应以及细胞分化抑制调节相关基因的表达在污染海域占据主导。
综上所述,各种假说的相关研究表明有机锡可能通过多种途径诱导性畸变,各假说之间在某些环节上可能存在交叉互作或者互补作用,进而形成Cross-talking,构建成复杂的信号传导网络,并且需要多个功能基因的参与。德国学者Stange等[74]提出狗岩螺性畸变涉及RXR和睾酮两种信号通路的参与,但目前似乎还没有在3种假说中建立某种联系(图1)。传统的单个或几个基因或蛋白的研究已经不可能阐明复杂的性畸变机制,而日益发展的高通量研究技术可以从全基因组尺度考察一个生理学过程,这为研究有机锡致海洋腹足类性畸变的分子机制和腹足类的防御应激机制提供了一条有效的途径。此外,众多的RXR参与海洋腹足类性畸变过程的证据表明,RXR在软体动物性别决定及性别分化过程中具有重要作用。相关的性畸变机制研究,揭示了软体动物性别决定及性别分化机制的特殊性,对该领域的进一步研究有可能突破人们对传统生殖系统发育理论的认识。
[1] 刘慧刚,徐立红.三丁基锡毒性作用生物标记研究进展[J].中华预防医学杂志,2005,39(4):288-289
[2] 陈天乙,郑素平,谭元生,等.有机锡化合物对水生无脊椎动物的毒性研究[J].环境化学,1994,13(3):266-268 Chen T Y,Zheng SP,Tan Y S,et al.A study on the toxicity of organotin on aquatic invertebrates system[J].Environmental Chemistry,1994,13(3):266-268(in Chinese)
[3] 孙红文,黄国兰,李书霞,等.三苯基锡和三丁基锡对大型蚤(Daphnia magna)的毒性作用研究[J].环境化学,2000,19(3):235-239 Sun H W,Huang G L,Li S X,et al.Study on toxic effects of and tributyltin on Daphniamagna[J].Environmental Chemistry,2000,19(3):235-239(in Chinese)
[4] 施华宏,黄长江.有机锡污染与海产腹足类性畸变[J].生态学报,2001,21(10):1711-1717 Shi H H,Huang C J.Tributyltin(TBT)pollution and imposex in marine gastropod[J].Acta Ecologica Sinica,2001,21(10):1711-1717(in Chinese)
[5] Sumpter JP,Johson A C.Lessons from endocrine disruption and their application to other issues concerning trace organics in the aquatic environment[J].Environmental Science &Technology,2005,39(12):4321-4332
[6] Gibbs PE,Bryan GW.Reproductive failure in populations of the dog-whelk,Nucella lapillus,caused by imposex induced by tributyltin from antifouling paints[J].Journal of the Marine Biological Association of the United Kingdom,1986,66(4):767-777
[7] Shi H H,Huang C J,Zhu SX,et al.Generalized system of imposex and reproductive failure in female gastropods of coastal waters of mainland China[J].Marine Ecology Progress Series,2005,304:179-189
[8] Huang C J,Zhu SX,Lin J,etal.Imposex ofMauritia arabica on the south-eastern coastof China[J].Journal of the Marine Biological Association of the United Kingdom,2008,88(7):1451-1457
[9] 曹伏君,罗杰,李长玲,等.细角螺的生殖系统组织学研究[J].热带海洋学报,2010,29(6):57-64 Cao F J,Luo J,LiC L,etal.Study on the reproductive system of Hemifusus ternatanus[J].Journal of Tropical Oceanography,2010,29(6):57-64(in Chinese)
[10] Sternberg R M,Gooding M P,Hotchkiss A K,et al.Environmental-endocrine control of reproductivematuration in gastropods:Implications for the mechanism of tributyltin-induced imposex in prosobranchs[J].Ecotoxicology,2010,19(1):14-23
[11] Horiguchi T,Lee JH,Park JC,et al.Specific accumulation of organotin compounds in tissues of the rock shell,Thais clavigera [J].Marine Environmental Research,2012,76:56-62
[12] Gibbs P E,Bryan GW,Pascoe P L,et al.The use of the dog-whelk,Nucella lapillus,as an indicator of tributyltin contamination[J].Journal of the Marine Biological Association of the United Kingdom,1987,67(3):507-523
[13] 施华宏,黄长江,雷瓒.利用波部东风螺(Babylonia formosae habei)性畸变现象监测海洋有机锡污染[J].海洋技术,2003,22(1):82-86 Shi H H,Huang C J,Lei Z.Organotin compound pollution in coastalwater as indicated by imposex in Babylonia formosae habei[J].Ocean Technology,2003,22(1):82-86(in Chinese)
[14] 汤丽,刘青坡,施华宏,等.厦门海域岩相海岸带疣荔枝螺(Thais clavigera)与甲虫螺(Cantharus cecillei)性畸变[J].生态学报,2009,29(9):4640-4648 Tang L,Liu Q P,Shi H H,et al.Imposex of Thais clavigera and Cantharus cecillei in the coastal waters of Xiamen[J].Acta Ecologica Sinica,2009,29(9):4640-4648(in Chinese)
[15] 黄晓丹,吴海莲,沈青,等.舟山海域腹足类动物性畸变研究[J].安徽农业科学,2011,39(22):13277-13278 Huang X D,Wu H L,Shen Q,etal.Studies on imposex of marine gastropods in Zhoushan[J].Journal of Anhui Agricultural Sciences,2011,39(22):13277-13278(in Chinese)
[16] Wu JY,Meng P J,Liu L L,etal.A high incidence of imposex in Pomacea apple snails in Taiwan:A decade after triphenyltin was banned [J].Zoological Studies,2010,49(1):85-93
[17] Mohamat-Yusuff F,Zulkifli SZ,Ismail A,etal.Imposex in Thais gradata as a biomarker for TBT contamination on the southern coast of Peninsular Malaysia [J].Water,Air& Soil Pollution,2010,211(1):443-457
[18] Castro I B,Fillmann G.High tributyltin and imposex levels in the commercial muricid Thais chocolata from two Peruvian harbor areas[J].Environmental Toxicology and Chemistry in Latin America,2012,31(5):955-960
[19] Horiguchi T,Kojima M,Hamada F,et al.Impact of tributyltin and triphenyltin on ivory shell(Babylonia japonica)populations[J].Environmental Health Perspectives,2006,114(1):13-19
[20] 施华宏,朱小兰,王蕾,等.腹足类性畸变研究进展[J].海洋环境科学,2009,28(4):463-468 Shi H H,Zhu X L,Wang L,et al.Advances of study on imposex in gastropods[J].Marine Environmental Science,2009,28(4):463-468(in Chinese)
[21] Castro L F C,Santos M M,Reis-Henriques M A.The genomic environmentaround the aromatase gene:Evolutionary insights[J].BMC Evolutionary Biology,2005,43(5):1-13
[22] Arai T,Harino H,OhjiM,etal.Ecotoxicology of Antifouling Biocides[M].Tykyo:Springer,2009:113
[23] Scott A P.Domollusks use vertebrate sex steroids as reproductive hormones?Part I:Critical appraisal of the evidence for the presence,biosynthesis and uptake of steroids[J].Steroids,2012,77(13):1450-1468
[24] Scott A P.Domollusks use vertebrate sex steroids as reproductive hormones?Part II:Critical review of the evidence that steroids have biological effects[J].Steroids,2013,78(2):268-281
[25] Guellec D L,Thiard M C,Adessi G L,et al.In vitro metabolism of androstenedione and identification of endogenous steroids in Helix aspersa[J].General and Comparative Endocrinology,1987,66(3):425-433
[26] Reis-Henriques M A,Guellec D L,Remy-Martin J P,et al.Studies of endogenous steroids from the marine mollusc Mytilus edulis by gas chromatography and mass spectrometry[J].Comparative Biochemistry and Physiology Part B:Comparative Biochemistry,1990,95(2):303-309
[27] Santos M M,Castro L FC,Reis-Henriques M A,etal.New insights into themechanism of imposex induction in the dogwhelk Nucella lapillus[J].Comparative Biochemistry and Physiology Part C:Toxicology&Pharmacology,2005,141(1):101-109
[28] Janer G,Lyssimachou A,Porte C,et al.Sexual dimorphism in esterified steroid levels in the gastropod Marisa cornuarietis:The effect of xenoandrogenic compounds[J].Steroids,2006,71(6):435-444
[29] Oberd rster E,McClellan-Green P.The neuropeptide APGWamide induces imposex in themud snail,Ilyanassa obsoleta[J].Peptides,2000,21(9):1323-1330
[30] Tillmann M,Schulte-Oehlmann U,Duft M,et al.Effects of endocrine disruptors on prosobranch snails(Mollusca:Gastropoda)in the laboratory.Part III:Cyproterone acetate and vinclozolin as antiandrogens[J].Ecotoxicology,2001,10(6):373-388
[31] Oehlmann J,Schulte-Oehlmann U,Bachmann J,et al.Bisphenol A induces superfeminization in the ramshorn snail Marisa cornuarietis(Gastropoda:Prosobranchia)atenvironmentally relevant concentrations[J].Environmental Health Perspectives,2006,114(s1):127-133
[32] Spooner N,Gibbs PE,Bryan GW,et al.The effect of tributyltin upon steroid titres in the female dogwhelk,Nucella lapillus,and the development of imposex [J].Marine Environmental Research,1991,32(1-4):37-49
[33] Bettin C,Oehlmann J,Stroben E.TBT-induced imposex inmarine neogastropods ismediated by an increasing androgen level[J].Helgoland Marine Research,1996,50(3):299-317
[34] Castro L FC,Melo C,SantosM M,etal.The estrogen receptor of the gastropod Nucella lapillus:Modulation following exposure to an estrogenic effluent?[J].A-quatic Toxicology,2007,84(4):465-468
[35] Jobling S,Casey D,Tyler C R,et al.Comparative responses ofmolluscs and fish to environmental estrogens and an estrogenic effluent[J].Aquatic Toxicology,2004,66(2):207-222
[36] 邓瑞鹏,韩雅莉,周小朋,等.雌雄及性畸变疣荔枝螺P450芳香化酶差异的研究[J].水生生物学报,2005,29(1):110-112 Deng R P,Han Y L,Zhou X P,et al.A study on the differences of cytochrome P450 aromatase in female,male and imposex samples of Thais clavigeras[J].Acta Hydrobiologica Sinica,2005,29(1):110-112(in Chinese)
[37] Morcillo Y,Ronis J,Porte M JC.Effects of tributyltin on the Phase I testosteronemetabolism and steroid titres of the clam Ruditapes decussata[J].Aquatic Toxicolo-gy,1998,42(1):1-13
[38] Ronis M J J,Mason A Z.The metabolism of testosterone by the periwinkle(Littorina littorea)in vitro and in vivo:Effects of tributyl tin[J].Marine Environmental Research,1996,42(1-4):161-166
[39] Gooding M P,LeBlanc G A.Biotransformation and disposition of testosterone in the easternmud snail Ilyanassa obsoleta[J].General and Comparative Endocrinology,2001,122(2):172-180
[40] Janer G,Sternberg R M,LeBlanc G A,et al.Testosterone conjugating activities in invertebrates:Are they targets for endocrine disruptors?[J].Aquatic Toxicology,2005,71(3):273-282
[41] Gooding M P,Wilson V S,LeBlanc G A,etal.The biocide tributyltin reduces the accumulation of testosterone as fatty acid esters in themud snail(Ilyanassa obsoleta)[J].Environmental Health Perspectives,2003,111(4):426-430
[42] Abidli S,Santos M M,Lahbib Y,et al.Tributyltin(TBT)effects on Hexaplex trunculus and Bolinus brandaris(Gastropoda:Muricidae):Imposex induction and sex hormone levels insights[J].Ecological Indicators,2012,13(4):13-21
[43] Kajiwara M,Kuraku S,Kurokawa T,etal.Tissue preferential expression of estrogen receptor gene in the marine snail,Thais clavigera [J].General and Comparative Endocrinology,2006,148(3):315-326
[44] Sternberg RM,Hotchkiss A K,LeBlanc G A.Synchronized expression of retinoid X receptor mRNA with reproductive tract recrudescence in an imposex-susceptible mollusk [J].Environment Science & Technology,2008,42(4):1345-1351
[45] Oberdorster E,Romano J,McClellan-Green P.The neuropeptide APGWamide as a penismorphogenic factor(PMF)in gastropod mollusks[J].Integrative and Comparative Biology,2005,45(1):28-32
[46] Féral C,LeGall S.The influence of a pollutant factor(tributyltin)on the neuroendocrinemechanism responsible for the occurrence of a penis in the female of Ocenebra erinaceain[C]//Lever J,Boer H H.Molluscan Neuro-Endocrinology.Proceedings of the International Minisymposium on Molluscan Endocrinology,North Holland,Amsterdam,1983:173-175
[47] Bryan GW,Bright D A.Uptake,tissue distribution and metabolism of 14C-labelled tributyltin(TBT)in the dog-whelk,Nucella lapillus[J].Journal of the Marine Biological Association of the United Kingdom,1993,73(4):889-912
[48] Horiguchi T,Kojima M,Kaya M,etal.Tributyltin and triphenyltin induce spermatogenesis in ovary of female abalone,Haliotis gigantea[J].Marine Environmental Research,2002,54(3-5):679-684
[49] Santos M M,Reis-Henriques M A,Sole M,et al.Triphenyltin and tributyltin,single and in combination,promote imposex in the gastropod Bolinus brandaris[J].Ecotoxicology and Environmental Safety,2006,64(2):155-162
[50] Castro L F C,Lima D,Machado A,et al.Imposex induction ismediated through the retinoid X receptor signalling pathway in the neogastropod Nucella lapillus[J].Aquatic Toxicology,2007,85(1):57-66
[51] Albalat R,Cañestro C.Identification of Aldh1a,Cyp26 and RAR orthologs in protostomes pushes back the retinoic acid geneticmachinery in evolutionary time to the bilaterian ancestor[J].Chemico-biological Interactions,2009,178(1-3):188-196
[52] Nishikawa J,Mamiya S,Horiguchi T,et al.Involvement of the retinoid X receptor in the development of imposex caused by organotins in gastropods[J].Environment Science& Technology,2004,38(23):6271-6276
[53] Bouton D,Escriva H,Pierce R J,et al.A conserved retinoid X receptor(RXR)from themollusk Biomphalaria glabrata transactivates transcription in the presence of retinoids[J].Journal of Molecular Endocrinology,2005,34(2):567-582
[54] Horiguchi T,Nishikawa T,Ohta Y,et al.Retinoid X receptor gene expression and protein content in tissues of the rock shell Thais clavigera[J].Aquatic Toxicology,2007,84(3):379-388
[55] Horiguchi T,Nishikawa T,Ohta Y,et al.Time course of expression of the retinoid X receptor gene and induction of imposex in the rock shell,Thais clavigera,exposed to triphenyltin chloride[J].Analytical and Bioanalytical Chemistry,2010,396(2):597-607
[56] Urushitani H,Katsu Y,Horiguchi T,et al.Cloning and characterization of retinoid X receptor(RXR)isoforms in the rock shell,Thais clavigera [J].Aquatic Toxicology,2011,103(1-2):101-111
[57] 张纪亮,朱文文,熊建利,等.有机锡通过核受体干扰内分泌的研究进展[J].环境科学导刊,2011,30(5):4-6 Zhang JL,Zhu W W,Xiong JL,et al.Review on endocrine disrupting effects of organotins via nuclear receptor[J].Environmental Science Survey,2011,30(5):4-6(in Chinese)
[58] Lima D,Reis-Henriques M A,Santos M M,et al.Tributyltin-induced imposex in marine gastropods in-volves tissue-specificmodulation of the retinoid X receptor[J].Aquatic Toxicology,2011,101(1):221-227
[59] Gooding M P.The role of steroid metabolism in tributyltin-induced imposex in the eastern mud snail(Ilyanassa obsoleta)[D].Raleigh:North Carolina State University,2002:214
[60] 李张伟,韩雅莉,简如君.性畸变疣荔枝螺体内保护酶系统活力的研究[J].海洋环境科学,2006,25(4):36-38 Li ZW,Han Y L,Jian R J.Study on protection ofactivity of endogenous enzymes system in imposex sample of Thais clavigera [J].Marine Environmental Science,2006,25(4):36-38(in Chinese)
[61] 李张伟,韩雅莉,李平,等.性畸变疣荔枝螺体内总蛋白和酯酶同工酶的比较研究[J].海洋环境科学,2008,27(5):470-474 Li ZW,Han Y L,Li P,et al.Studies on total protein and esterase isoenzyme of imposex Thais clavigera [J].Marine Environmental Science,2008,27(5):470-474(in Chinese)
[62] 徐小妹,许孙曲.丁基锡化合物的物种形成和遗传毒性[J].国外锡工业,1995,23(4):53-57
[63] Dopp E,Hartmann L M,Rettenmeier AW,et al.The cyto-and genotoxicity of organotin compounds is dependent on the cellular uptake capability [J].Toxicology,2007,232(3):226-234
[64] Liu H G,Wang Y,Lian L J,et al.Tributyltin induces DNA damage as well as oxidative damage in rats[J].Environmental Toxicology,2006,21(2):166-171
[65] Zuo ZH,Wang CG,Wu M F,et al.Exposure to tributyltin and triphenyltin induces DNA damage and alters nucleotide excision repair gene transcription in Sebastiscusmarmoratus liver[J].Aquatic Toxicology,2012,122-123:106-112
[66] 管云雁,韩雅莉.两种荔枝螺染色体核型以及性畸变个体染色体研究[J].海洋环境科学,2004,23(3):21-23 Guan Y Y,Han Y L.Studies on karyotype of two species of Thais and variations of chromosome from imposex individual[J].Marine Environmental Science,2004,23(3):21-23(in Chinese)
[67] 周小朋,韩雅莉,何薇薇.RAPD技术在疣荔枝螺性畸变机理方面研究中的应用[J].海洋环境科学,2004,23(4):31-34 Zhou X P,Han Y L,He W W.Application of RAPD on the study of imposexmechanism in Thais clavigera[J].Marine Environmental Science,2004,23(4):31-34(in Chinese)
[68] Hagger J A,Depledge M H,Galloway T S,et al.Is there a causal association between genotoxicity and the imposex effect[J].Environmental Health Perspectives,2006,114(s1):20-26
[69] Iguchi T,Watanabe H,Katsu Y.Application of ecotoxicogenomics for studying endocrine disruption in vertebrates and invertebrates[J].Environmental Health Perspectives,2006,114(s1):101-105
[70] 贾锡伟,张子平,邹志华,等.三丁基锡暴露条件下杂色鲍肝胰腺均一化cDNA文库的构建[J].生物技术通报,2009(11):136-139 Jia X W,Zhang Z P,Zou Z H,et al.Construction of normalized cDNA library from hepatopancreas of abalone Haliotis diversicolor supertexta exposure to tributyltin[J].Biotechnology Bulletin,2009(11):136-139(in Chinese)
[71] Jia XW,Zou ZH,Wang G D,et al.Gene expression profiling in respond to TBT exposure in small abalone Haliotis diversicolor[J].Fish Shellfish Immunology,2011,31(4):557-563
[72] Pascoal S,Carvalho G,Vasieva O,etal.Transcriptomics and in vivo tests reveal novelmechanisms underlying endocrine disruption in an ecological sentinel,Nucella lapillus[J].Molecular Ecology,2013,22(6):1589-1608
[73] Titley-O'Neal C P,Spade D J,Zhang Y P,et al.Gene expression profiling in the ovary of Queen conch(Strombus gigas)exposed to environments with high tributyltin in the British Virgin Islands[J].Science of the Total Environment,2013,449(1):52-62
[74] Stange D,Sieratowicz A,Oehlmann J.Imposex development in Nucella lapillus-Evidence for the involvement of retinoid X receptor and androgen signalling pathways in vivo[J].Aquatic Toxicology,2012,106-107:20-24 ◆
