高浓度发酵乳基料的制备及复合酶对其特性的影响
2013-08-07鲍志宁林伟锋
鲍志宁,林伟锋*,叶 君,熊 犍*
(华南理工大学轻工与食品学院,广东 广州 510640)
目前,乳酸菌发酵乳制品种类繁多,在各国社会经济和日常饮食中都起着重要作用[1-3]。高浓度发酵乳基料是一种新型的发酵乳制品,其蛋白质含量为6%~10%,而普通发酵乳中蛋白质含量为2.9%~3.5%[4]。高浓度发酵乳基料具有蛋白质含量高、氨基酸含量高和酸度高的特点,其作为一种创新型发酵乳基料将逐渐引起食品行业的关注。将发酵乳作为乳基料添加至其他食品中,如加至发酵乳饮料[5-6]和烘焙制品所用的香基[7-8]中也是发酵乳基的新型用途。
目前在奶味香基领域中,脂肪酶酶解乳脂制作香基已被广泛应用[9]。但酶法所得香基香型单调,易呈酸败味,因此将微生物发酵与酶法结合是制备高品质天然发酵乳香基料的重要途径。现有研究中二者结合的研究主要为先发酵后酶解。于铁妹等[10]将1.6%全脂乳经德氏乳杆菌发酵后再加入脂肪酶酶解获得天然奶味香基料。Tsai等[11]在乳酸菌发酵5h后,结合蛋白酶酶解来去除蛋白质含量约为4%的发酵豆乳中的苦味并使其富含生物活性肽和γ-氨基丁酸(GABA)。而将乳酸菌发酵与酶解同步结合的研究很少,二者同步作用制备的高浓度发酵乳基料具有更为突出的发酵特征,将对发酵乳基料的研究和应用具有积极意义。
本实验通过高浓度发酵乳基料及与复合酶制剂酶解同步结合,来改善和提高其酸度和氨基酸态氮含量等特征,以期为高浓度发酵乳基料的应用研究,提供新的实验依据和参考。
1 材料与方法
1.1 材料与试剂
全脂奶粉(批号9121477) 雀巢公司;无水奶油(批号1233456) 新西兰恒天然公司;干酪乳杆菌(Lactobacillus casei,LC-L134-1-P) 本实验室保存;复合酶(脂肪酶与蛋白酶符合,脂肪酶活力15000U/L,蛋白酶酶活力1.4AU/L) 广州华琪生物科技有限公司;其他试剂均为分析纯。
1.2 仪器与设备
PHS-25型pH计 上海精密科学仪器有限公司;SZX超净工作台 上海浦东跃新科学仪器厂;PYXDHS·400-BS恒温培养箱 北京市医疗设备厂。
1.3 方法
1.3.1 高浓度发酵乳基的制作
1.3.1.1 高浓度发酵乳基的配方
全脂奶粉30%(蛋白质含量7.8%)、无水奶油5%、水70%、菌种干酪乳杆菌0.4‰。
1.3.1.2 高浓度发酵乳基的工艺
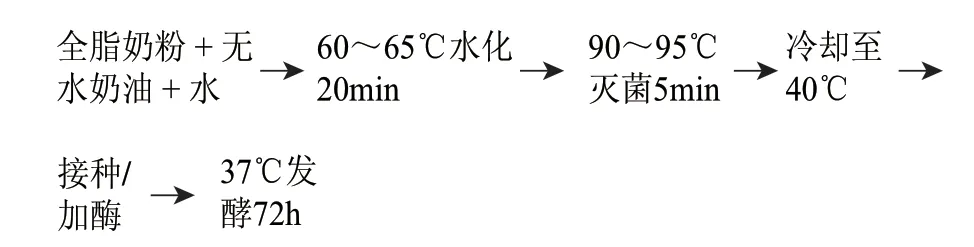
1.3.2 复合酶对高浓度发酵乳基的影响
1.3.2.1 复合酶的添加对高浓度发酵乳基的影响
在接种/加酶时,1#样仅接种干酪乳杆菌发酵;2#样为仅添加复合酶酶解;3#样为干酪乳杆菌发酵与复合酶酶解相结合。对比3种条件下高浓度发酵乳基发酵/酶解过程产物的变化。
1.3.2.2 不同的复合酶添加量对高浓度发酵乳基的影响
在接种/加酶时,分别添加0.1‰(4#样)、0.2‰(5#样)、0.3‰复合酶(6#样)与乳酸菌发酵相结合,对比不同复合酶添加量对高浓度发酵乳基发酵过程的影响。
1.3.3 高浓度发酵乳基发酵过程中指标的测定
pH值的测定采用PHS-25型酸度计测定。酸度的测定采用°T表示发酵乳基的酸度[12]。活菌数的测定参见文献[13]。氨基酸态氮含量的测定参见文献[12]。
2 结果与分析
2.1 高浓度发酵乳基料发酵过程的发酵特性变化
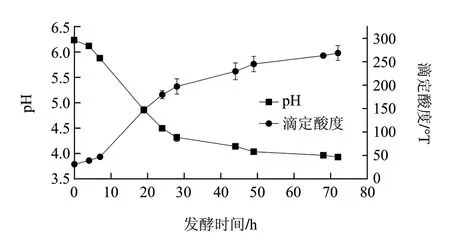
图 1 高浓度发酵乳基酸度随发酵时间的变化Fig.1 Time course of pH and titratable acidity during fermentation
由图1可知,随着发酵时间的延长,pH值下降,下降速率由低到高再降低,发酵72h时pH值降至3.93。而普通发酵乳(蛋白质含量3.0%,奶油含量0%)在发酵72h时pH值为3.6~3.7[14]。同时,随着发酵时间的延长,高浓度发酵乳基料的滴定酸度上升,上升速率变化也与pH值下降速率变化相对应,发酵72h时滴定酸度为268.5°T。普通的发酵乳发酵72h时滴定酸度仅为125~135°T。由于高浓发酵乳基料具有较高的蛋白质含量和脂肪含量,对于干酪乳杆菌产生的乳酸有较强的缓冲作用,因此即使滴定酸度很高,pH值也仅为4.0左右。高浓发酵乳基料的滴定酸度是普通发酵乳的2倍,若应用于发酵乳饮料调配,添加量仅需20%~30%即可达到适宜的酸度。
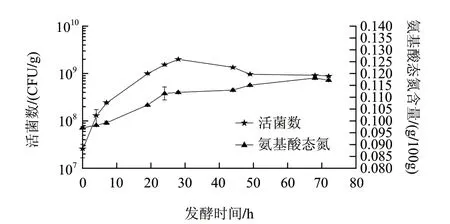
图 2 高浓度发酵乳基活菌数和氨基酸态氮随发酵时间的变化Fig.2 Time course of the viable cell number of Lactobacillus casei and amino nitrogen content during fermentation
由图2可知,干酪乳杆菌在发酵28h时由对数期进入稳定期,与图1中产酸速率变化拐点一致。同时,在发酵28~72h过程中,活菌数稳定在1×109CFU/mL,这一方面是因为高浓度发酵乳基中含有大量的碳源和氮源,另一方面说明该干酪乳杆菌具有对较高酸度具有耐受性的特征[15]。图2中氨基酸态氮含量在发酵过程中的变化呈现缓慢波动上升的趋势。乳酸菌在增殖过程中,不断地分泌蛋白酶水解乳蛋白产生游离氨基酸,产生的游离氨基酸不仅用于满足干酪乳杆菌对氮源的需要,而且可以作为发酵风味物质[16]。发酵72h后,高浓度发酵乳基中的氨基酸态氮含量由初始的0.097g/100g增加至0.117g/100g,提高了20.6%。而同样菌株发酵72h,普通发酵酸奶的氨基酸态氮含量仅为0.061g/100g。
2.2 复合酶对高浓度发酵乳基料发酵的影响
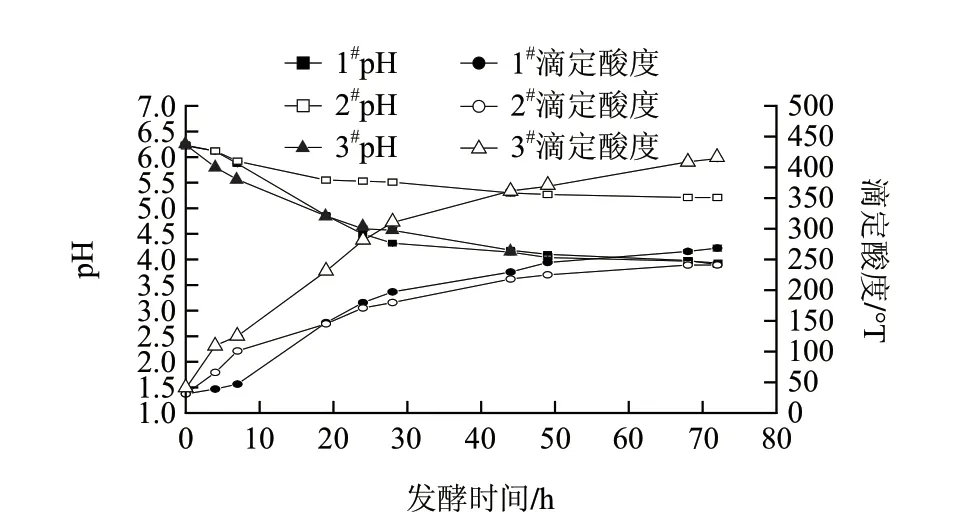
图 3 复合酶对高浓度发酵乳基酸度的影响Fig.3 Time course of pH and titratable acidity during simultaneous hydrilysis and fermentation
由图3可知,仅添加复合酶的高浓度发酵乳基料(2#样)pH值下降幅度比添加了干酪乳杆菌的高浓度发酵乳基料(1#样和3#样)小,发酵72 h时,2#样pH值为5.21,而1#样和3#样pH值均为3.90,且二者在发酵过程中pH值变化趋势基本一致。同时乳酸菌与复合酶相结合的高浓度发酵乳基料(3#样)的滴定酸度远高于1#样和2#样,近于2#样滴定酸度的两倍。发酵72 h时,3#样滴定酸度为414.4°T, 2#样仅为241.1°T。这是由于复合酶中的脂肪酶会将乳脂分解成脂肪酸,脂肪酸的羧基使乳基料具有较高的滴定酸度和pH值。3#样中由于复合酶中的蛋白酶酶解乳蛋白,为乳酸菌提供了更多氨基酸作为速效氮源,促进了菌株的增殖和乳酸分泌,进而与脂肪酸协同获得更高的滴定酸度。
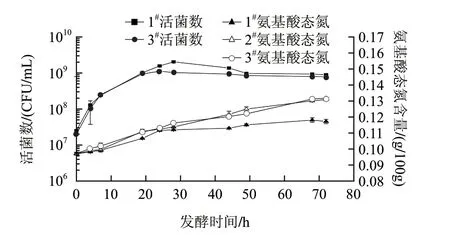
图 4 复合酶对高浓度发酵乳基活菌数和氨基酸态氮含量的影响Fig.4 Time course of the viable cell number of Lactobacillus casei and amino nitrogen content during simultaneous hydrilysis and fermentation
2#样品在发酵过程中无菌落检出。由图4可知,3#样中干酪乳杆菌于20h时从对数期进入稳定期,1#样则于28h转折。同时在发酵50h之前,1#样的活菌数均比3#样高。这是因为乳酸菌内生性酶构成简单,许多氨基酸不能由自身合成,因此乳酸菌培养时往往需要含有丰富氨基酸的物质来提供氮源[17-18]。蛋白酶分解的氨基酸作为速效氮源促进菌株生长繁殖,但氮源过多则会使菌体生长过于旺盛,反而加速菌体的衰老和自溶[19-20],菌株分裂与衰退均加速,因此活菌数没有明显增加。随着酶解脂肪酸的积累使滴定酸度大幅升高,也在一定程度上抑制了干酪乳杆菌的生长。因此3#样在发酵20~72h活菌数保持平稳。2#样和3#样的氨基酸态氮含量随着酶解和发酵时间的延长不断增加,并且比1#样有大幅提高。在72h处,氨基酸态氮含量1#样为0.117g/100g,2#样为0.131g/100g,3#样为0.132g/100g,3#样比1#样提高12.8%。
因此,乳酸菌发酵剂与酶制剂复配将发酵和酶解同步进行获得的高浓度发酵乳基料具有更高的酸度和氨基酸态氮含量,使其作为一种发酵乳基料的特征更为突出,更有利于其在各类食品中的应用。
2.3 不同复合酶添加量对高浓度发酵乳基料发酵的影响
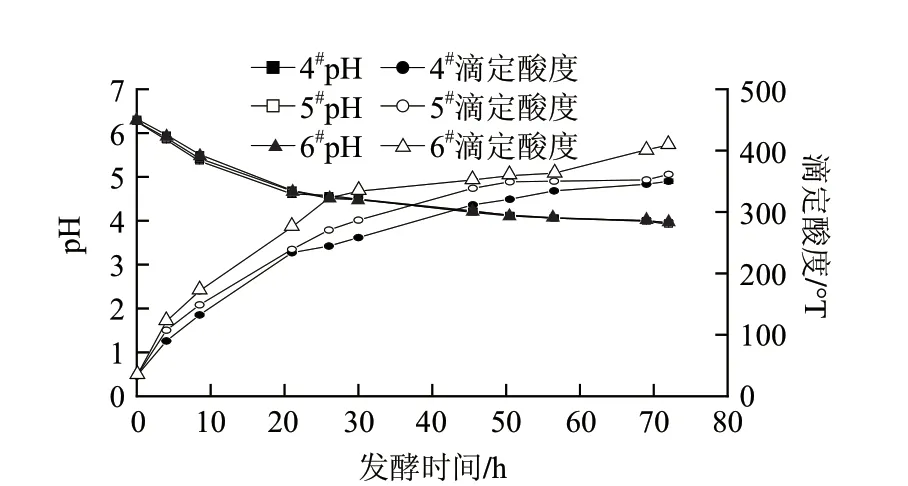
图 5 不同复合酶添加量对高浓度发酵乳基酸度的影响Fig.5 Effect of enzyme dosage on pH and titratable acidity
由图5可知,3种不同浓度的复合酶添加量(4#样0.1‰,5#样0.2‰,6#样0.3‰)的高浓度发酵乳基料pH值变化趋势和数值是完全一致的,最终pH值在3.92~3.98之间。而滴定酸度则受复合酶添加量的影响明显,复合酶添加量高的高浓度发酵乳基料滴定酸度也相应较高,由高到低依次为409.2、361.2、350.1°T,同时3个样品的滴定酸度变化趋势是基本一致。因此可以选择不同的复合酶添加量和发酵时间来满足不同的产品需求。
由图6可知,3个样品在活菌数变化趋势和数值上是完全一致的,这可能是因为复合酶中蛋白酶酶解乳蛋白产生氨基酸的量远远超过干酪乳杆菌增殖需要,以及当酸度超过某个值其对干酪乳杆菌的抑制程度也趋向一致。高浓度发酵乳基料的氨基酸态氮含量随着复合酶添加量的增加在发酵后期(46h之后)有不同程度的提高,4#样为0.125g/100g,6#样为0.134g/100g。3个样品氨基酸态氮含量的变化趋势也与滴定酸度保持一致。
3 结 论
本实验研究了蛋白质含量为7.8%的高浓度发酵乳基料在发酵过程中的动力学参数变化情况,获得的高浓度发酵乳基料的滴定酸度和氨基酸态氮含量均较高,同时高浓度缓冲体系使pH值也较高,干酪乳杆菌LC-L134-1-P对酸的良好耐受性使其活菌数稳定在1×109CFU/mL左右。本实验进一步将复合酶制剂与乳酸菌发酵剂复配获得的高浓度发酵乳基料具有更高的酸度和氨基酸态氮含量。同时复合酶含量越高,高浓度发酵乳基料的酸度和氨基酸态氮含量越高。综合看来,经过发酵和酶解的高浓度发酵乳基料具有更高的酸度和氨基酸态氮含量的特征,接下来将对其在发酵过程中酶与发酵剂同步作用机理,以及高浓度发酵乳基料在风味、在焙烤等食品中的增香应用和功能性做更深入研究。
[1] NARVHAS J A, OSTERAAS K, MUTUKUMIRA T, et al. Production of fermented milk using a malty compound-producing strain of Lactobacillus lactis subsp. latis biovar. diacetylactis isolated from Zimbabwean naturally fermented milk[J]. International Journal of Food Microbiology, 1998, 41: 73-80.
[2] GONFA A, FOSTER H A, HOLZAPFEL W H. Field survey and literature review on traditional fermented milk products of Ethiopia[J]. International Journal of Food Microbiology, 2001, 68: 173-186.
[3] 张和平. 益生乳酸菌及发酵乳的开发趋势[J]. 农产品加工, 2012(6): 8-9.
[4] 中华人民共和国卫生部.GB19302—2010 发酵乳[S]. 北京: 中国标准出版社, 2010.
[5] 徐致远, 杭锋, 郭翔, 等. 影响褐色乳饮料中益生菌数量的因素[J]. 食品科学, 2010, 31(13): 193-195.
[6] 郭翔, 徐致远, 艾连中, 等. 褐色益生菌乳饮料稳定性研究及工艺条件优化[J]. 食品科学, 2009, 30(22): 226-230.
[7] 李慧东, 刘冠勇. 用乳酸菌和酵母共同发酵法生产面包的研究[J]. 食品工业, 2007(6): 25-26.
[8] 汪薇, 赵文红, 白卫东, 等. 乳酸菌发酵制备天然奶味香精的研究[J]. 食品与发酵工业, 2010, 36(10): 191-195.
[9] 于铁妹, 徐迅, 王永华, 等. 脂肪酶水解乳脂制备天然奶味香基的研究[J]. 食品与发酵工业, 2008, 34(4): 150-153.
[10] 于铁妹, 王永华, 杨博, 等. 微生物发酵与酶法结合制备天然奶味香基的研究[J]. 食品与发酵工业, 2008, 34(10): 153-157.
[11] TSAI J S, LIN Y S, PAN B S, et al. Antihypertensive peptides and γ-aminobutyric acid from prozyme 6 facilitated lactic acid bacteria fermentation of soymilk[J]. Process Biochemistry, 2006, 41: 1282-1288.
[12] 张水华. 食品分析[M]. 北京: 中国轻工业出版社, 2004: 79-174.
[13] 凌代文. 乳酸菌分类鉴定及实验方法[M]. 北京: 中国轻工业出版社, 1999: 85-86.
[14] ARYANA K J, MCGREW P. Quality attributes of yogurt with Lactobacillus casei and various prebiotics[J]. LWT-Food Science and Technology, 2007, 40(10): 1808-1814.
[15] 杜昭平, 马成杰, 孙克杰, 等. 干酪乳杆菌LC2W发酵脱脂乳营养因子优化[J]. 食品科学, 2012, 33(19): 205-210.
[16] KRANENBURG R V, KLEEREBEZEM M, VLIEG J V H, et al. Flavour formation from amino acid by lactic acid bacteria predictions from genome sequence analysis[J]. International Dairy Journal, 2002, 12: 111-121.
[17] HUJANEN M, LINKO S, LINKO Y Y, et al. Optimization of media and cultivation conditions for L(+)(S)-lactic acid production by Lactobacillus casei NNRL B-441[J]. Journal of Applied Microbiology and Biotechnology, 2001, 56(1/2): 126-130.
[18] HUJANEN M, LINKO Y Y. Effect of temperature and various nitrogen sources on L(+)-lactic acid production by Lactobacillus casei[J]. Journal of Applied Microbiology and Biotechnology, 1996, 45(6): 307-313.
[19] 曹军卫, 马辉文. 微生物工程[M]. 北京: 科学出版社, 2002: 80-88.
[20] 余龙江. 发酵工程原理与技术应用[M]. 北京: 化学工业出版社, 2006: 89-93.
