黄海南部和东海小黄鱼资源动态的比较
2013-07-07刘尊雷谢汉阳严利平袁兴伟杨林林黎雨轩吴颖程家骅
刘尊雷,谢汉阳,严利平,袁兴伟,杨林林,黎雨轩,吴颖,程家骅
(1.中国水产科学研究院东海水产研究所,农业部东海与远洋渔业资源开发利用重点实验室,上海200090;2.浙江象山县海洋与渔业局,浙江宁波315700;3.唐山市科学技术情报研究所,河北唐山063000)
黄海南部和东海小黄鱼资源动态的比较
刘尊雷1,谢汉阳2,严利平1,袁兴伟1,杨林林1,黎雨轩1,吴颖3,程家骅1
(1.中国水产科学研究院东海水产研究所,农业部东海与远洋渔业资源开发利用重点实验室,上海200090;2.浙江象山县海洋与渔业局,浙江宁波315700;3.唐山市科学技术情报研究所,河北唐山063000)
根据2006—2007年底拖网调查资料,利用体长频率法估算了黄海南部和东海小黄鱼Larimichthys polyactis种群的生长、死亡参数,比较了两海域小黄鱼资源变动的空间特征差异,并利用动态综合模型分析了小黄鱼资源的利用状况。结果表明:黄海南部和东海小黄鱼种群渔获年龄主要为0~1+a,以当龄鱼(0 a)为主;平均极限体长(L∞)分别为226.47、257.25 mm,平均生长速度(K)分别为0.39/a、0.43/a,理论生长起点年龄(t0)分别为-0.80 a、-0.31 a;总死亡系数(Z)分别为1.81/a、3.05/a,捕捞死亡系数(F)分别为1.42/a、2.62/a,东海种群的捕捞强度大于黄海南部种群。渔获概率分析显示:黄海南部种群渔获概率为25%、50%和75%的选择体长分别为103.6、112.4、121.2 mm,东海种群分别为110.5、122.4、130.5 mm,东海种群的开捕体长稍大于黄海南部种群。动态综合模式分析表明,黄海南部种群资源利用参考点E0.1(单位补充量渔获量曲线初始斜率为10%时的开发率)、E50%(资源量为初始资源量50%时的对应开发率)、Emax(最大产量时的对应开发率)分别为0.560、0.386和0.649,东海种群分别为0.563、0.356和0.669,当前开发率已经超过了目标参考点E0.1,资源处于过度利用状态。
小黄鱼;生长;死亡;渔获概率;Beverton-Horl模型
小黄鱼 Larimichthys polyactis为暖温性底层鱼类,广泛分布于中国的东海、黄海、渤海和朝鲜的西部海域[1-2]。20世纪60年代至80年代,小黄鱼的捕捞强度不断增大,小黄鱼资源逐渐衰退。近年来,东海区实施伏季休渔制度,小黄鱼补充群体得到有效的养护,资源量有所恢复[3],但渔获物主要以当龄鱼(0 a)为主,个体呈小型化和低龄化,性成熟提前,资源基础依然薄弱[4]。
目前,中国对小黄鱼种群划分仍存在较大争议。胡建成[5]、林新濯等[6]根据小黄鱼的地理分布和形态特征,以32°N为分界线,将东海区小黄鱼分为两个种群,即黄海南部种群和东海种群;徐兆礼等[7]则认为,这两个种群在越冬场存在混栖现象,两者实为同一种群。小黄鱼的分布特征是对栖息环境长期适应的结果,不同群系鱼类的生理活动和对外部的调控能力不同,长期的环境变化、生态过程和应对捕捞压力的响应导致种群生活史参数和种群动态变化也不同。认识鱼类种群的空间结构有助于提高渔业资源评估和管理能力[8]。鉴于小黄鱼种群认定的模糊性,本研究中采用胡建成[5]、林新濯等[6]的种群划分结果,即假定东海区小黄鱼分为黄海南部种群和东海种群,拟用2006年1月至2007年12月小黄鱼时间序列长度频率数据,估算独立种群的生长参数、死亡参数和渔获概率,建立综合动态模式,评价黄海南部种群和东海种群的空间资源利用状况,分析不同群系小黄鱼的最适开捕标准,旨在为管理部门的决策和小黄鱼资源的合理利用提供科学依据。
1 材料与方法
1.1 研究海域
研究海域为东海中部至黄海南部区域(27°00'N~34°30'N,122°00'E~127°00'E),沿岸至大陆架边缘覆盖水深20~125 m。依据脊椎骨、背鳍、臀鳍、鳃耙、幽门盲囊、膘支管等6项分节特征的种群群系划分方法[6],选取32°00'N~34°00'N和123°00'E~126°00'E为黄海南部小黄鱼种群栖息水域(主要为黄海南部和东海北部部分区域,包括吕四渔场、大沙渔场中南部、沙外渔场西部海域),31°30'N以南为东海小黄鱼种群栖息水域(包括长江口渔场、江外渔场和以南各渔场)(图1)。
1.2 方法
1.2.1 数据来源 生物学测定数据来源于2006—2007年东海区底拖网渔业资源调查和常规监测,其中调查按春夏秋冬(4、6、9、12月)4个季节进行,其他月份的样品则通过监测渔船的取样进行补充。调查船为205.07 kW底拖网渔船,网口为96 m×4 m,网囊网目为20 mm,平均拖速为2~3 n mile/h。
在实验室测定渔获样品的体长(精确到 1 mm)和体质量(精确到0.1 g),对黄海南部种群和东海种群小黄鱼分别测定了2 341尾和2 586尾。1.2.2 种群的生长参数 假定小黄鱼生长过程可用非季节性Von Bertalanffy生长方程拟合,利用Fi-SATⅡ软件分析每月的体长频率数据。首先通过修正的Powell-Wetheral法(以较高体长组渔获进行拟合,不受捕捞网具选择性能的影响),结合Beverton-Holt死亡系数计算方程对预处理的数据进行线性拟合,计算公式为

其中:L'为每个组距的下限值;L∞为生长的渐进体长;Z为总死亡系数;K为平均生长速率。由此可估算出L∞=-a/b,Z/K=-(1+b)/b。然后通过Elefan依次重新构建体长频率数据,并以上述计算的L∞作为初始值,对Von Bertalanffy生长方程进行拟合,数次迭代后获得最优生长参数K和L∞,以score值作为拟合度评价指标,其值分布在0与1之间。
1.2.3 种群的死亡参数 用渔获量曲线求得总死亡系数。渔获量曲线的线性方程为
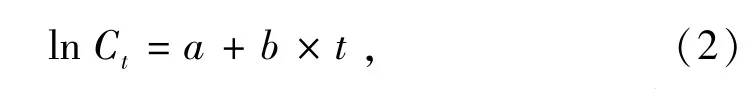
其中:Ct为t龄时的渔获数量;a、b为待估参数,且b为总死亡系数Z。渔获量曲线随相对年龄变化趋势越陡峭,死亡系数就越高。由于样本的年龄未能鉴定,因此,采取变换体长渔获量曲线来估算总死亡系数Z,修正后的方程为
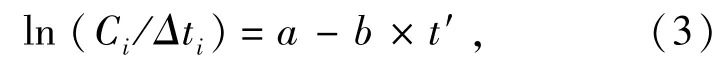
其中:Ci为第i体长组渔获尾数;t'为第i体长组中值体长对应的年龄,即相对年龄;Δti为第i体长组从下限体长生长至上限体长的时间。
用Pauly公式计算自然死亡系数M[9]:
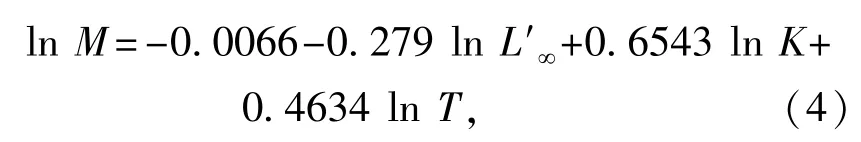
其中:L'∞为渐近全长(cm);K为平均生长速率; T为栖息环境的年平均水温。本研究中,黄海南部和东海种群栖息环境的年平均水温分别为13.6[10]、20℃[11]。
1.2.4 渔获概率 选择捕获概率为50%时的对应体长作为动态综合模型中的开捕体长,通过渔获量曲线左支可以近似得到该值。拖网渔具的选择曲线为逻辑斯谛曲线,将其转化为线性方程为
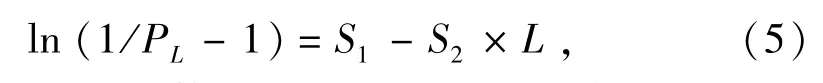
其中:PL为对应体长为L时的捕获概率;S1和S2为待估参数。L0.50=S1/S2,为渔获概率为50%时的选择体长。
1.2.5 渔业管理参考点 用Beverton-Holt近似模型计算单位补充量的相对渔获量和相对生物量[12]。假设同一世代鱼类在相同年龄通过刀刃型补充曲线各自达到补充年龄(tr)、开捕年龄(tc)和从种群消失时的最大年龄(tmax),因此模型更适合于生命周期较长种类,其表达式分别为
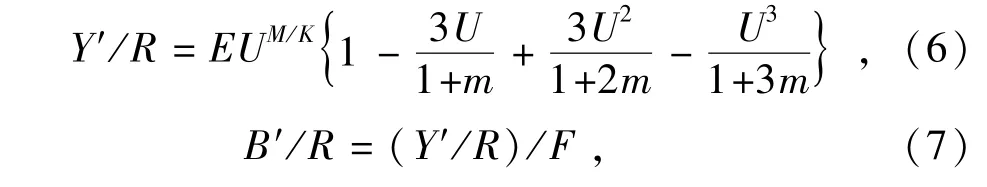
其中:R为补充量;B'为相对生物量;Y'为相对渔获量;F为捕捞死亡系数;Y'/R为单位补充量相对渔获量;B'/R为单位补充量相对生物量;E为开发率;U=1-(Lc/L∞),为生长到开捕体长以上的群体可被渔业利用的比例,U值取决于Lc,Lc为渔获概率为50%时对应的选择体长;m=K/Z。
分别以E0.1(单位补充量渔获量曲线初始斜率为10%时的开发率)、E50%(资源量为初始资源量50%时的对应开发率)、Emax(最大产量时的对应开发率)作为统计指标来衡量渔业的开发程度,并改变开发率,评价单位补充量相对渔获量和单位补充量相对生物量的变化,探索资源合理利用的参考点。
2 结果与讨论
2.1 种群的生长参数
2.1.1 群体的年龄组成 假设黄海南部和东海小黄鱼种群的体长结构均为正态分布,采用最大似然估计法获得群体的年龄组成结果如图2所示。两个种群均由0 a和1+a结构组成,其中黄海南部种群0 a、1+a平均体长分别为113.61、151.31 mm,0 a、1+a群体的比例分别占79.8%、20.2%;东海种群0 a、1+a平均体长分别为 128.62、149.2 mm,0 a、1+a群体的比例分别占 82.8%、17.2%。
2.1.2 生长参数估算 以Elefan技术重建体长频率直方图,估计生长参数。由于该方法忽略了生长起点年龄,根据Bhattacharya分割的0 a组和1+a组平均体长,结合已计算的参数K和L∞,取均值可估算出理论生长起点年龄(t0),黄海南部种群和东海种群的t0分别为-0.80 a、-0.31 a。生长方程为
黄海南部种群Lt=226.47×(1-e-0.39×(t+0.80)),
东海种群Lt=257.25×(1-e-0.43×(t+0.31))。

图2 黄海南部和东海种群的年龄-体长分布频数Fig.2 The age-length frequency of small yellow croaker populations in Southern Yellow Sea and East China Sea
2.2 种群的死亡参数
采用变换体长渔获量曲线的下降右支估算总死亡系数(图3),其中黄海南部种群的总死亡系数为1.81/a(95%置信区间为1.68/a~1.94/a),东海种群总死亡系数相对较高,为3.05/a(95%置信区间为2.66/a~3.44/a)。黄海南部和东海种群的自然死亡系数分别为0.39/a和0.48/a,捕捞死亡系数分别为1.42/a和2.62/a,当前开发率分别为0.78和0.86。表明,东海种群的利用强度较高。

图3 采用变换体长渔获量曲线估计小黄鱼种群的总死亡系数Fig.3 Total mortality estimated by length-converted catch curves in small yellow croaker populations
2.3 渔获概率
渔获概率通过变换体长渔获量曲线的上升左支部分求得(图4),即每个体长组的实际样本数和期望样本数的比值,以累积渔获概率为50%时所对应的中值体长作为动态综合模型中的开捕体长Lc。黄海南部小黄鱼种群渔获概率为25%、50%和 75% 时的选择体长 L0.25、 L0.50、 L0.75分别为103.6、112.4、121.2 mm,东海种群则分别为110.5、122.4、130.5 mm。

图4 渔获概率为25%、50%和75%时的选择体长Fig.4 The estimation of selection body length for L0.25,L0.5,and L0.75of small yellow croaker populations
2.4 单位补充量的相对渔获量和相对生物量
单位补充量的相对生物量(B'/R)结果(图5)表明,E0.1作为目标参考点比Emax更为合理。黄海南部小黄鱼种群开发率为E0.1时,B'/R约下降至初始资源量的30%;开发率为Emax时,B'/R已下降至初始资源量的25%以下,而单位补充量的相对渔获量(Y'/R)并没有明显增加。东海种群同样如此。故本研究中选取E0.1作为资源评价与管理的目标参考点。黄海南部和东海小黄鱼种群的E0.1分别为0.560和0.563,当前的资源开发率分别为0.78和0.86,已经超过了参考点,B'/R也已经降到极低水平,表明资源已经处于过度利用状态,如果不采取管理措施,亲体量减少到一定程度会导致资源衰竭而不可恢复。

图5 单位补充量的相对渔获量(Y'/R)和相对生物量(B'/R)Fig.5 Relative yield per recruit and relative biomass per recruit for small yellow croaker populations using the knife edge selection procedure
3 讨论
3.1 小黄鱼种群资源的空间差异
对小黄鱼生长参数的分析显示,黄海南部小黄鱼生长速度慢,渐进体长小,生态寿命长,这是对栖息生态环境长期适应的结果。随着纬度的增加,水温降低,鱼类的寿命延长,生长率减小[13]。在捕捞压力相对较小的环境中,鱼生长缓慢,生殖群体的平均年龄和最大年龄均提高。小黄鱼在适宜环境下具有生长缓慢以及世代性成熟持续时间较长的特点,因此,小黄鱼种群比其他生命周期短的种群稳定性更大,生态适应性更小[1]。当捕捞强度破坏了种群原有的稳定性时,小黄鱼通过增大个体生长速率、性成熟提前等调节机制以促进种群数量的恢复,适应新的栖息环境。不同海区鱼类的种群密度、饵料基础等特定生态环境之间的差异也决定了这种生长性能的差异,但在实际计算过程中,生长参数L∞、t0和K起着互相修正的作用,黄海南部和东海小黄鱼种群生长参数不同,不能通过单个参数分析各海区的生长差异。林龙山[4]采用生长性能指数从多元角度比较了黄海南部和东海小黄鱼种群,认为后者的生长性能指数略大于前者。
3.2 渔获概率差异分析
在补充曲线和网目选择性的共同影响下,期望尾数和实际捕获尾数的差值趋向最小。本研究中利用体长变换渔获量曲线估算了未完全补充群体的捕获概率为50%时的选择体长,该方法的前提是各个年龄组的总死亡系数相同。实际上,对于网目选择性较低的年龄组,捕捞死亡系数要小于渔获物完全留存阶段;而幼鱼的自然死亡系数通常远高于高年龄组,因此,各年龄组总死亡系数只能近似认为是常数,这种影响很少引起关注。
小黄鱼补充群体的体长结构呈正态分布,其进入渔场的阶段便不相同,补充群体随体长变化的补充量曲线也表现为逻辑斯谛曲线形式[14]。利用上述方法得到的选择性曲线是补充曲线和网目选择性曲线的合成曲线。当网目尺寸充分大时,补充曲线和网目选择性曲线的体长范围不存在相互交错,这时可以忽略补充过程,合成曲线即为网目选择性曲线;而当网目尺寸过小,补充曲线和网目选择性曲线的体长范围出现重合,此时得到的50%选择性指标即为合成曲线所对应的相对体长,并不能代表真实网具的选择性能(实际选择性曲线介于合成曲线和补充曲线之间)。本研究中计算的黄海南部和东海小黄鱼种群渔获概率为50%时的选择体长(即开捕体长)分别为112.4、122.4 mm,大于网具选择性所对应的体长,导致动态综合模型采用的参数值Lc/L∞过大,低估了小黄鱼资源的实际利用状态。黄海南部和东海种群的高开发率水平,进一步证实了小黄鱼渔业资源的过度开发。
3.3 复合种群对渔业管理的意义
实行科学管理,控制鱼类资源衰竭,需要明确管理对象的种群结构。种群数量的变动取决于补充、生长、捕捞死亡和自然死亡,而捕捞量、年龄组成等生物学指标,可初步阐述种群的数量、结构和兴衰状况[15]。在渔业资源评估和管理中,忽视种群的空间结构可导致渔业资源被过度捕捞,如北大西洋鳕Gadus morhua[16]和大西洋鲱Clupea harengus[8]。现行的渔业管理方法,将小黄鱼群体视作单一的种群结构,忽略了亚种群的空间异质性[17]。小黄鱼种群划分并无明确定论,但根据两个独立种群的假设,东海种群小黄鱼遭受更高的开发率,远远超出渔业管理的参考点水平,而捕捞压力需要以平均生产力为尺度,这时确定性的渔业管理可能会制定错误的管理目标,导致地方种群的衰退。Ying等[18]提出真实种群、孤立种群和随机交流种群3种情况,模拟了生物量和渔获量的变化轨迹,认为采用单一种群管理方案,所有的亚种群均会被过度捕捞,陷入种群枯竭。由于该文的模型结构未考虑到物种的生活史,忽视了不同种群的自身变化规律,如遗传多样性、生长率、死亡率、繁殖力和应对外部环境干扰的调控能力等,研究结果略显悲观,但在一定程度上促进了渔业管理策略的优化。
在独立种群的情况下,降低捕捞强度尽管能使资源利用趋于合理,但根据Beverton-Holt模型研究结果,降低捕捞强度的作用并不明显,综合考虑社会、经济因素,缩减捕捞强度执行难度较大。而提高开捕年龄可以取得显著效果,但这种效果因种群结构而不同,张国政等[10]认为,在维持捕捞死亡系数1.63不变的情况下,黄海南部小黄鱼开捕年龄应提高到1.41 a,单位补充量的渔获量将比现行渔业点增加45.26%。林龙山等[17]将黄海南部和东海小黄鱼种群作为独立种群进行分析,提出小黄鱼的最适开捕年龄为2.10 a。本研究结果表明,东海种群和黄海南部种群当前的开捕年龄(L0.5对应的年龄)分别为1.19 a和0.96 a,均低于上述最适开捕年龄。可见,若不考虑种群的空间结构,单独采用黄海南部种群的管理方案,小黄鱼渔业资源将面临较大的衰退风险。因此,在实际渔业管理中,应充分考虑不同种群的空间差异,调整捕捞努力量分配方案,提高渔业资源评估和管理能力。
小黄鱼可能由多个地方种群组成,种群间既相互独立,又有个体间基因交流。环境塑造、生殖洄游和越冬洄游等物理-生态固有特性在诸多方面限制了现有种群的识别与鉴定,使传统的渔业资源评估多基于混合种群或理想情景的分析,种群空间结构的研究也仅停留在科学理论的层面。行政区划和地理区域的渔业管理仍是当前资源养护的必不可少的方案,而基于遗传种群结构的渔业管理在渔业实践中尚无法开展,需进一步研究。
[1]赵传絪,陈永法,洪港船,等.东海区渔业资源调查和区划[M].上海:华东师范大学出版社,1987:339-356.
[2]陈红波,李继龙,杨文波,等.东黄海小黄鱼秋季索饵环境栖息指数的研究[J].大连海洋大学学报,2011,26(4):348-351.
[3]林龙山,程家骅.延长东海区伏季休渔期的渔业效果分析[J].大连水产学院学报,2009,24(1):12-16.
[4]林龙山.黄海南部和东海小黄鱼渔业生物学及其管理策略研究[D].青岛:中国海洋大学,2009.
[5]胡建成.从嵊泗县线外船捕捞结合黄海东海渔海况速报谈南黄海族小黄鱼的划分[J].海洋渔业,1998(1):29-35.
[6]林新濯,邓思明,黄正一.小黄鱼种群生物学测定学的研究[C]//海洋渔业资源论文选集.北京:农业出版社,1965.
[7]徐兆礼,陈佳杰.再议中国近海小黄鱼种群的划分问题[J].应用生态学报,2010,21(11):2856-2864.
[8]Stephenson R L.Stock complexity in fisheries management:a perspective of emerging issues related to population sub-units[J]. Fisheries Research,1999,43:247-249.
[9]Pauly D.On the interrelationships between natural mortality,growth parameters and mean environmental temperature in 175 fish stocks [J].ICES Journal of Marine Science,1980,39(2):175-192.
[10]张国政,李显森,金显仕,等.黄海南部小黄鱼生长、死亡和最适开捕体长[J].中国水产科学,2010,17(4):839-846.
[11]林龙山,程家骅.东海区小黄鱼渔业生物学现状的分析[J].中国海洋大学学报,2004,34(4):565-570.
[12]Pauly D,Soriano M L.Some practical extensions to Beverton and Holt's relative yield-per-recruit model[R].Manila:Asian Fisheries Society,1986.
[13]Gordon H C,Michael G F,Miroslaw P,et al.Life-time growth patterns of pumpkinseed Lepomis gibbosus introduced to Europe, relative to native North American populations[J].Folia Zoologica,2004,53(3):237-254.
[14]詹秉义.渔业资源评估[M].北京:中国农业出版社,1995: 43-45.
[15]刘文斌,张红林.鱼类种群研究在渔业管理中的应用[J].淡水渔业,1992(4):42-44.
[16]Hutchinson W F.The dangers of ignoring stock complexity in fishery management:the case of the North Sea cod[J].Biology Letters,2008(4):693-695.
[17]林龙山,程家骅,凌建忠,等.东海区主要经济鱼类开捕规格的初步研究[J].中国水产科学,2006,13(2):250-256.
[18]Ying Y P,Lin L S,Gao T X,et al.Risks of ignoring fish population spatial structure in fisheries management[J].Canadian Journal of Fisheries and Aquatic Sciences,2011,68(12):2101-2120.
Comparative population dynamics of small yellow croaker Larimichthys polyactis in Southern Yellow Sea and East China Sea
LIU Zun-lei1,XIE Han-yang2,YAN Li-ping1,YUAN Xing-wei1, YANG Lin-lin1,LI Yu-xuan1,WU Ying3,CHENG Jia-hua1
(1.Key Laboratory of East China Sea&Oceanic Fishery Resources Exploitation and Utilization,Ministry of Agriculture,East China Sea Fisheries Research Institute,Chinese Academy of Fishery Sciences,Shanghai 200090,China;2.Xiangshan Bureau of Ocean and Fisheries,Ningbo 315700, China;3.Institute of Scientific and Technical Information of Tangshan,Tangshan 063000,China)
Growth parameters(L∞and K)and mortality coefficients(Z,M and F)of small yellow croaker Larimichthys polyactis populations were estimated in the coastal waters of Southern Yellow Sea(SYS)and East China Sea(ECS)from January 2006 to December 2007 by length distribution data collected by trawl survey.The current exploitation status and rational utilization of the small yellow croaker resources were evaluated by target reference point calculated through modified Beverton-Holt dynamic pool model.The Bhattacharya's analysis revealed that the age composition of the small yellow croaker was 0 year and 1+year,dominated by the group of 0 year.Fitting the von Bertalanffy growth function to these data showed asymptotic length(L∞)of 226.47 mm and 257.25 mm,curvature parameter(K)of 0.39 and 0.43 per year,and initial condition parameter,(t0)of-0.80 a and-0.31 a for the SYS and ECS population,respectively.Total mortality coefficients was found to be 1.81 per year in SYS and 3.05 per year in ECS population,fishing mortality coefficients 1.42 per year in SYS population and 2.62 per year in ECS population,higher capture intensity in ECS population than that in the SYS population.The estimated length at different probability of capture was found to be L0.50=112.4 mm,L0.25=103.6 mm,and L0.75=121.2 mm in the SYS population and L0.50=122.4 mm,L0.25=110.5 mm,and L0.75=130.5 mm in ECS population,higher in the ECS than that in the SYS.Dynamic pool models showed that the small yellow croaker population in the SYS had prevailing E0.1(developing rate at 10%of the initial increment,0.560),E50%(developing rate at 50%of the initial increment,0.386)and Emax(developing rate at the maximal yield,0.649),and that the small yellow croaker population showed in E0.1of 0.563,E50%of 0.356 and Emaxof 0.669 in the ECS,indicating that the stock was overexploited due to above prevailing E0.1.
small yellow croaker;growth;mortality;probability of capture;Beverton-Horl model
S931
A
2013-03-10
国家自然科学基金青年基金资助项目(31101901);中央级公益性科研院所基本科研业务费专项基金资助项目(2009T06);农业部近海渔业资源监测调查专项(2006—2007);国家公益性行业(农业)科研专项(201303047)
刘尊雷(1982-),男,助理研究员。E-mail:liuzl@eastfishery.ac.cn
程家骅(1965-),男,研究员。E-mail:ziyuan@sh163.net
2095-1388(2013)06-0627-06
