5种植物上扶桑绵粉蚧的适生性及其潜在为害分析
2013-06-11王前进齐国君吕利华何余容
王前进,高 燕,陈 婷,齐国君,吕利华,何余容
(1.华南农业大学资源环境学院,广州 510642;2.广东省农业科学院植物保护研究所,广东省植物保护新技术重点实验室,广州 510640)
扶桑绵粉蚧Phenacoccus solenopsis Tinsley 隶属于半翅目Hemiptera,蚧总科Coccoidea(Tinsley,1898;Cockerell,1902),是我国的一种外来入侵新害虫(武三安,张润志,2009)。该虫1898年首次在美国新墨西哥州被发现(Tinsley,1898),后在中美洲、加勒比海地区、厄瓜多尔等地严重发生(Williams et al.,1992)。2002年以来,该虫相继入侵智利、阿根廷、巴西、印度、巴基斯坦和尼日利亚(Larrain,2002;Granara,2003;Culik and Gullan,2005;Akintola and Ande,2008)。2008年在我国广州市扶桑植物上首次发现该虫为害(武三安,张润志,2009),截至2010年分布至我国12 省市(自治区),对扶桑等园林绿化树造成了影响(武三安,张润志,2009;徐卫等,2009)。世界范围内,该害虫已对棉花种植造成了严重为害,2005年其入侵巴基斯坦、印度后,在棉花种植区暴发成灾,造成了巨大的经济损失(Dutt,2007;Hodgson,2008;Nagrare et al.,2009)。虽然该虫尚未对我国棉花产业造成严重危害,但其在中国的适生区较广,而且已在各主要棉花种植区已有零星分布,因此其对棉花种植区的潜在危害值得极大关注(王艳平等,2009;马骏等,2011)。
此外,扶桑绵粉蚧是一种多食性害虫,寄主范围极广。在巴基斯坦其寄主包括18科55种植物,主要包括棉花、扶桑、豚草、向日葵、南瓜、大戟、茄等(Hodgson,2008),后来其寄主种类又增加到53科154种,包括20种大田作物、45种园艺植物、64种杂草、25种灌木等,主要隶属锦葵科Malvaceae、菊科Asteraceae、茄科Solanaceae、苋科Amaranthaceae、大戟科Euphorbiaceae和葫芦科Cucurbitaceae(Arif et al.,2009)。在我国浙江省其寄主植物有33科66种(周湾等,2012)。在印度的棉区,银胶菊Parthenium hysterophorus、苍耳Xanthium stumarium 等杂草是扶桑绵粉蚧越冬的主要中间寄主,并为该粉蚧次年在棉田发生提供了充足的虫源基数(Saini et al.,2009)。
目前,国内外分别以棉花、土豆苗、番茄、茄子及南瓜等经济作物为寄主研究了该害虫的生长发育及种群增长(Vennila et al.,2010;Fand et al.,2010;黄芳等,2011;王莹莹等,2012),并比较了几种经济作物对扶桑绵粉蚧的适合性,为分析其对棉花等经济作物的潜在危害提供了依据。但是在扶桑绵粉蚧如此广泛的寄主范围内,除了经济作物以外,杂草也是重要的一部分,如豚草、蟛蜞菊等入侵性杂草(Hodgson,2008;Arif et al.,2009;周湾等,2012)。目前,尚无扶桑绵粉蚧在杂草上生长发育及适合性的研究报道。杂草上扶桑绵粉蚧生长发育与生殖的研究不仅会扩大其寄主适生性范围,而且会为明确入侵杂草在扶桑绵粉蚧周年发生与传播扩散中所起的作用提供科学依据。
表3中centrality是中介中心性,是测度节点在网络中重要性的一个指标。突发性探测可以看出关键词的词频在某一段时间有很大的变化,strength是突发强度,图5中有6个关键词在相应的起始时间段内词频变化明显,说明这一段时间内关注研究此关键词的文章比较密集。通过中介中心性指标和突发性探测可以观测出热点关键词和重要文献。通过统计出的2004年至2018年移动阅读研究关键词的词频、中介中心性及突发性可以归纳出移动阅读有以下几个研究方向比较受学者关注:移动图书馆、数字出版、阅读推广、电子阅读器、阅读行为、电子书、移动阅读服务等。
本研究拟在恒温条件下以菊科植物的豚草和三裂蟛蜞菊、锦葵科的棉花和扶桑及茄科植物番茄为供试寄主,观察比较五种植物对扶桑绵粉蚧生长发育、繁殖的影响,研究扶桑绵粉蚧对不同科植物的适生性,以分析入侵杂草在扶桑绵粉蚧扩散传播和周年转主为害中的潜在作用,同时也为分析该害虫对我国棉花生产的潜在为害及暴发成灾机制提供科学依据。
1.2 临床资料收集 收集所有患者入急诊科后的基础临床数据,包括病史、年龄、性别、心电图、高血压病、糖尿病、高脂血症等,所有病例入院当天急诊抽血查肌钙蛋白Ⅰ,分别运用 HEART评分和MEWS评分进行病情评估。
1 材料与方法
1.1 供试虫源及植物品种
供试虫源:以采自广东省广州市天河区内绿化带扶桑植株上的扶桑绵粉蚧为供试虫源。将采集的粉蚧饲养在(26~31)℃、(75±5)%和L∶D=14∶10 人工气候室中,以南瓜果实为食物,进行保种和继代饲养。
扶桑购于广东省广州市岭南花卉市场。
番茄品种为广东省农业科学院蔬菜研究所生产的皇冠666。
雌虫的产卵量测定表明,在豚草上的扶桑绵粉蚧每雌产卵量最高,为511.12 粒,与扶桑上的495.98 粒无显著性差异;在棉花上的粉蚧产卵量为463.73 粒,显著高于其取食三裂蟛蜞菊时的产卵量(434.98 粒);在番茄上的粉蚧雌虫产卵量为342.63±7.09 粒,均显著低于上述几种试验处理的,其产卵量较豚草和扶桑的产卵量下降32.9%和30.92%。
棉花品种为湘杂九号。
三裂蟛蜞菊、豚草幼苗采自广东省农业科学院大丰基地。
所有植物均种植在直径20 cm、高30 cm 的塑料花盆中,选取营养生长的健康植株作为供试植物。
1.2 试验方法
扶桑绵粉蚧若虫生长发育:选择产卵初期的雌成虫,将卵囊完全剔除,置于1个新鲜无虫的南瓜果实上,24 h 后从新形成卵囊内剥离出若虫,将其作为供试虫体。摘取植株心叶下第3 至第5叶位的叶片,并用湿润的棉花包裹叶柄保湿,再将叶片置于皿底铺有湿润滤纸的直径14 cm 塑料培养皿中,每皿1片叶片。培养皿盖上人工挖出直径4 cm 的孔洞,且用300目尼龙纱布密封。试验时,用3 号狼毫描笔(上海周虎臣)从卵囊内挑取1头初孵若虫接到供试植物叶片上,再将培养皿置于人工气候箱(28±1℃、75±5%、L∶D=14∶10)观察。处理虫数120头,每皿1头。每天7∶30和19∶30 观察,并记录供试虫的发育虫态和死亡情况。试验观察至供试若虫发育至成虫为止。1龄粉蚧若虫活动能力强,须用Parafilm(BEMIS)密封培养皿。每2~3 d更换1次叶片。移虫时,首先用柔软的毛笔尖轻轻触碰虫体,待其将口针从叶片中拔出,再将其转移到新鲜叶片上。每种植物处理的扶桑绵粉蚧若虫为120头。
“不出来就好了,比坐牢还惨一点。”李高明当然没坐过牢,手上的老茧、夜里的寒风,虽然难忍,还不至于让他这么想。最痛苦的是李高明回不了家。
自体输血也有禁忌证,以下患者不适合自体输血:采血可能诱发疾病加重者、菌血症患者、贫血、肝肾功能不良或严重心脏病等患者[20]。
五种植物上扶桑绵粉蚧的产卵历期略有差异,在豚草上粉蚧雌虫产卵历期为15.84 d,显著短于取食番茄、三裂蟛蜞菊、棉花、扶桑的雌虫产卵历期(分别为18.89 d、18.93 d、18.50 d和17.86 d),且这四种植物上的雌虫产卵历期无显著性差异。
1.3 数据分析
植物种类不同,成虫寿命也有较大差异(表1、2)。番茄上的扶桑绵粉蚧雌虫寿命最长,为50.55 d,显著地长于其它四种植物上的;三裂蟛蜞菊、棉花和扶桑上的雌虫寿命分别为48.14 d、46.73 d和45.88 d,三裂蟛蜞菊和扶桑有显著差异,棉花则与两者差异不大;相应的若虫期发育历期最短,成虫阶段的产卵前期和产卵期都最短,使得豚草上的雌虫寿命最短,为41.79 d。不同植物上雄虫的寿命与雌虫寿命有相似的趋势。雄虫寿命在番茄上为23.59 d,显著长于其它四种植物上的;其次是三裂蟛蜞菊上的,为21.15 d;再次是棉花和扶桑上的雄虫寿命,分别为19.41 d、18.23 d,且两者间无显著差异;雄虫寿命最短的为豚草上的扶桑绵粉蚧种群,为16.75 d,显著短于另外四种植物上的雄虫寿命。
结果表明,在豚草上扶桑绵粉蚧的雌虫和雄虫的发育最快,发育历期最短,寿命最短;但在番茄上其发育最慢,发育历期最长,寿命最长。在锦葵科的棉花和扶桑上饲喂的雌虫和雄虫的历期及寿命均无显著性差异,但饲喂在菊科植物豚草和三裂蟛蜞菊上的雌雄虫历期及寿命差异较大,前者均显著高于后者。
2 结果与分析
2.1 五种植物对扶桑绵粉蚧雌虫存活率的影响
统计了取食番茄、三裂蟛蜞菊、棉花、扶桑和豚草的扶桑绵粉蚧雌虫在各发育阶段的年龄特征存活率(图1)。由于在1龄若虫阶段死亡的虫体无法区分性别,故将其既归入雌虫,也归入雄虫。
番茄上的扶桑绵粉蚧产卵前期最长,为12.28 d,显著长于其他四种植物上扶桑绵粉蚧的产卵前期;豚草上的扶桑绵粉蚧产卵前期最短,为10.05 d,显著地短于三裂蟛蜞菊、棉花、扶桑上的粉蚧(分别为11.58 d、11.01 d、11.34 d),其中后三种植物上的粉蚧产卵前期无显著性差异。

图1 五种植物对扶桑绵粉蚧雌虫存活率的影响Fig.1 Effects of five plants on survival rate of female mealybug
2.2 五种植物上扶桑绵粉蚧发育历期及寿命
在实验室条件下,测定了取食番茄、三裂蟛蜞菊、棉花、扶桑和豚草植株的扶桑绵粉蚧雌虫的生长发育历期(表1)。就扶桑绵粉蚧雌虫而言,在五种植物上的生长发育历期是不同的。在番茄上,雌虫1龄若虫的发育历期为7.50 d,显著地长于其它四种植物上饲喂的扶桑绵粉蚧的发育历期;在其余四种植物中,在豚草上其发育历期最短,为5.74 d,但其在三裂蟛蜞菊、棉花、扶桑间的差异不显著。取食三裂蟛蜞菊的2龄若虫的发育历期最长,为4.61 d,显著长于取食扶桑和豚草的粉蚧发育历期,而取食其它两种植物-三裂蟛蜞菊、棉花的粉蚧发育无显著差异。取食番茄的3龄若虫的发育历期最长,达5.72 d,显著地长于在豚草上的粉蚧发育历期4.91 d,但取食其他三种植物的3龄若虫间无显著性差异。取食番茄的粉蚧雌成虫历期最长,为32.73 d,其次依次为三裂蟛蜞菊、棉花、扶桑,取食豚草的雌成虫发育历期最短,为27.53 d。
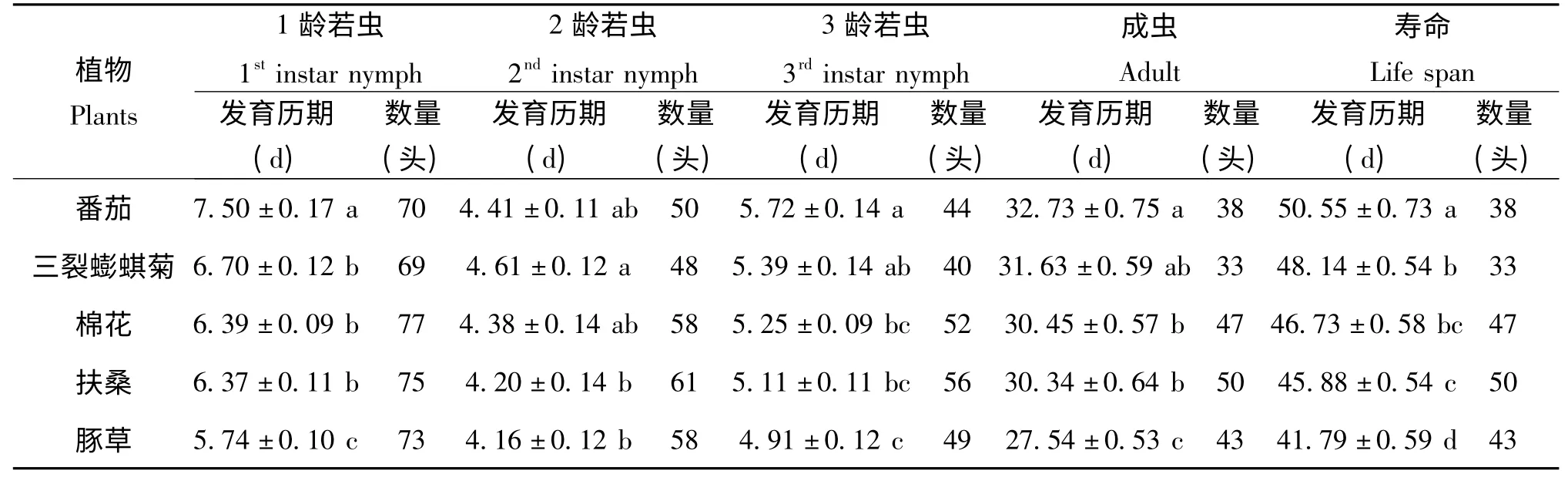
表1 扶桑绵粉蚧雌虫取食不同植物的发育历期及寿命Table 1 Developmental duration and life span of female mealybug on different plants
在五种植物上生长发育的扶桑绵粉蚧雄虫的发育历期也存在差异。1龄若虫的发育历期在番茄上最长(6.97 d),显著长于三裂蟛蜞菊、棉花、扶桑上的,分别为6.35 d、5.85 d、5.94 d,但三者间无显著性差异,且又显著长于豚草上的发育历期(5.34 d)。2龄若虫取食番茄时历期最长,为6.58 d,显著长于其余四种供试植物饲喂的扶桑绵粉蚧的历期;在后四种中,三裂蟛蜞菊上的为5.67 d,棉花、扶桑和豚草上的扶桑绵粉蚧的发育历期分别为4.92 d、4.70 d、4.58 d,而且后三者间无显著性差异。番茄和豚草上扶桑绵粉蚧蛹期历期为最长和最短的,分别为6.32 d和5.38 d,而饲喂在其它3种植物上的粉蚧蛹期长度无差异。番茄和三裂蟛蜞菊上的扶桑绵粉蚧成虫历期较长,分别为3.09 d和2.92 d,两者间无显著性差异,其次为棉花、扶桑、豚草上的成虫历期,分别为2.41 d,2.31 d,1.89 d;棉花和豚草间的差异性显著,扶桑与它们均无显著性差异。
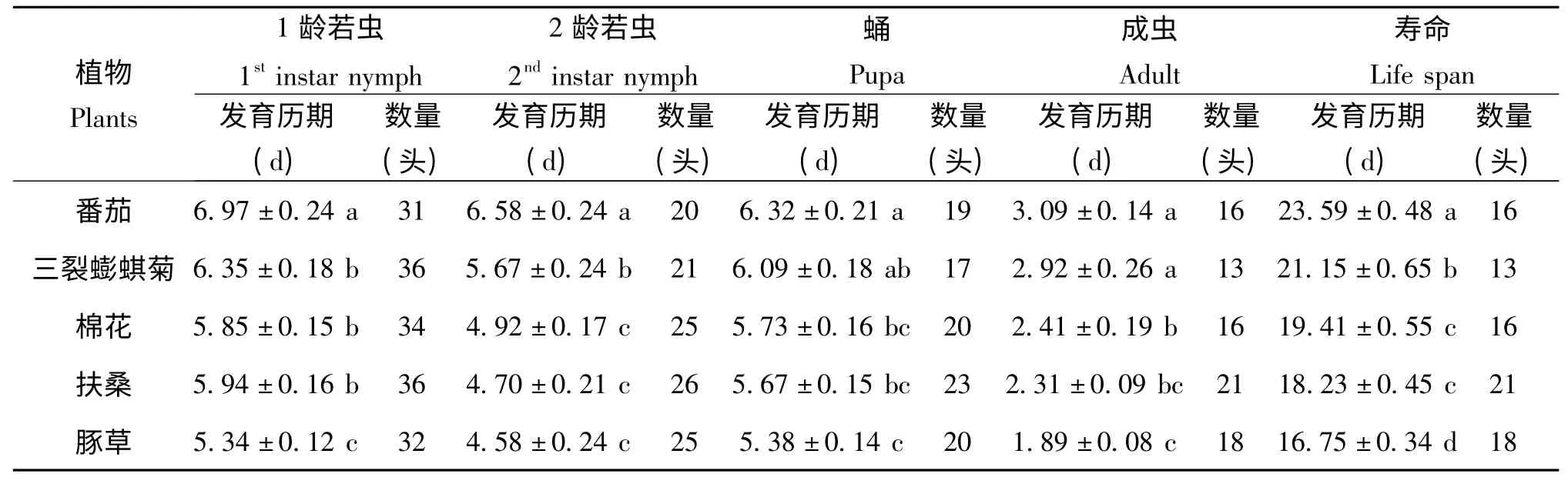
表2 扶桑绵粉蚧雄虫取食不同植物的发育历期及寿命Table 1 Developmental duration and life span of male mealybug on different plants
根据五种植物上扶桑绵粉蚧的存活率、雌虫产卵量及发育历期,计算扶桑绵粉蚧实验种群增长参数。种群增长参数的计算参照徐汝梅(1987):净增殖率R0=∑lxmx;世代平均周期T=∑lxmx/ R0;内禀增长率rm=(ln R0)/T;周限增长率λ=erm;种群加倍时间t=ln2/ rm。其中,x 是发育时间,lx是任一个体在x 期间存活的概率;mx表示x 期间平均每雌产卵数。
采用DPS 数据处理系统,对试验数据进行Duncan's 新复极差法进行多重比较(唐启义,冯明光,2002)。
由于人字形波纹板片具有传热性能好、承压能力大等优点,现有板式换热器板片波纹形状以人字形最多,大多数的研究也主要针对人字形波纹。德国人Focke是首位研究“通过改变人字形板式换热器板片波纹倾角等参数提高换热效率”的学者。Mehrabian等通过截取最小计算区域及采用计算流体动力学模拟研究了板式换热器,研究表明板片波纹形状对换热和压降有着十分重要的影响。曲宁对板式换热器的流动及传热特性进行数值研究,结果表明波纹倾角、波纹深度、波纹间距等参数对板式换热器的流场及温度场分布有重要影响。
2.3 不同植物上扶桑绵粉蚧雌虫的生殖参数
当扶桑绵粉蚧若虫羽化成成虫后,即取雌雄虫配对,获得雌成虫生殖参数(表3)及日产卵量变化(图2)。
假设方位向间歇采样周期Tsa=2T,采样占空比Da=0.5,即每间隔一个SAR脉冲才采样转发一次.图3为a、b通道接收干扰信号的状态,实心圆点代表接收信号中含有干扰信号,如n和n+2时刻;空心圆点代表无干扰信号,如n+1和n+3时刻.方位向间歇采样导致相邻时刻总是不能同时接收到干扰信号,此时对区域目标进行分析,结合式(1)可得b通道n、n+1时刻和a通道n+1、n+2时刻接收的信号为
结果表明:取食不同种植物的扶桑绵粉蚧存活率存在差异,其中取食三裂蟛蜞菊的虫体存活率最低,其1龄、2龄、3龄若虫和成虫的存活率分别为87.50%、69.57%、57.97%和47.83%,其次为番茄,其1龄、2龄、3龄若虫和产卵前期的存活率分别为84.17%、71.43%、62.86%和54.29%;豚草和棉花上相对应的参数分别为87.50%、79.45%、67.12%、58.90%和92.50%、75.32%、67.53%、61.04%;扶桑上该粉蚧的存活率最高,其1龄、2龄、3龄的若虫和产卵前期的存活率分别为92.50%、81.33%、74.67%和66.67%,平均存活率为78.79%。数据表明锦葵科的扶桑和棉花上扶桑绵粉蚧存活率较大,较适宜于扶桑绵粉蚧存活;菊科的豚草上扶桑绵粉蚧平均存活率也较大,但在同属于菊科的三裂蟛蜞菊上扶桑绵粉蚧则表现出最小的存活率,是本试验五种植物中最不适宜其存活的植物。
扶桑绵粉蚧成虫的寿命及繁殖力测定:当上述试验中的扶桑绵粉蚧发育为成虫后,将取同种植物上的雌虫、雄虫各1头放入培养皿中配对,逐日观察记录雌虫的产卵起始时间、每日产卵量,至雌虫死亡为止。计算出产卵前期、产卵期、产卵量,从而得出各处理的雌成虫寿命及繁殖力。每种处理观察33~50 组。
原来,几年前,蜡像馆生意火爆,李总不顾牟月身体虚弱,强迫他们夫妻俩加班赶工,导致牟月病情加重,最后不幸身亡。而如今,李总又要坚持拆除蜡像馆。眼见自己和妻子一生的心血即将毁于一旦,陆影气愤难平,最终走上歧途,在这个雪夜刺杀了李总。

图2 五种植物上扶桑绵粉蚧的日产卵量变化规律Fig.2 Regular pattern of egg number of a day per female on different plants
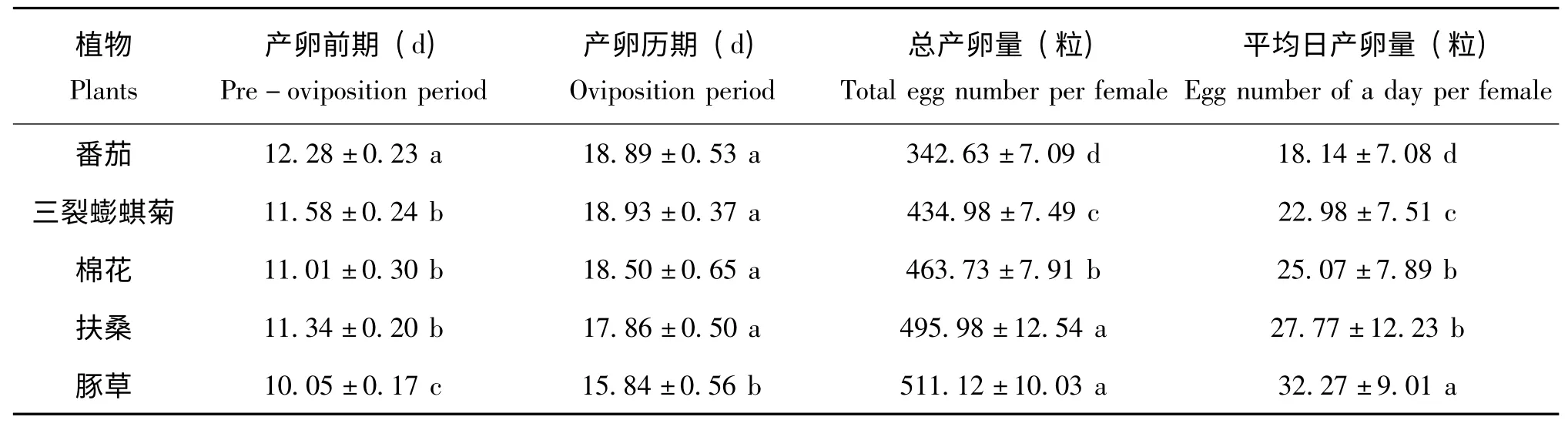
表3 五种植物上扶桑绵粉蚧雌虫的生殖参数Table 3 Reproductive parameter of female mealybug on different plants
由于取食不同植物的扶桑绵粉蚧雌虫发育速率不同,五种植物上扶桑绵粉蚧雌成虫进入产卵期的时间略有差异,先后顺序为豚草、扶桑、棉花、三裂蟛蜞菊和番茄,而且扶桑绵粉蚧每雌日平均产卵量也表现出一定趋势(图1)。豚草上其为32.27 粒,为最大,其次是扶桑,27.77 粒,再次是棉花,25.07 粒,其后为三裂蟛蜞菊、番茄,其每雌日平均产卵量约为22.98 粒和18.14 粒。
2.4 五种植物上扶桑绵粉蚧的实验种群参数
依据室内饲养的观察结果,组建了扶桑绵粉蚧在五种植物上的实验种群繁殖特征生命表,计算出净增殖率R0、平均世代周期T、内禀增长率rm、周限增长率λ和种群加倍时间t 等实验种群参数(表4)。结果表明:扶桑上的净增殖率R0最大,为283.79,在豚草上扶桑绵粉蚧净增殖率R0次之(274.33),内禀增长率rm(0.18)与周限增长率λ(1.20)最大,平均世代周期T、种群加倍时间t 最短(31.44、3.88);在番茄上R0、rm、λ值最小,分别为161.41、0.13、1.14,T和t 最长,为38.38和5.23。研究表明,豚草和扶桑是供试植物中最适合扶桑绵粉蚧的生长发育、繁殖的植物,棉花、三裂蟛蜞菊次之,番茄为最不适合的植物。
3 结论与讨论
本试验主要研究了扶桑绵粉蚧在五种植物上的生长发育及繁殖,比较了这五种植物上的种群参数,内禀增长率rm,研究结果表明扶桑绵粉蚧种群呈明显增长趋势,种群增长能力强,表明五种供试植物是扶桑绵粉蚧的寄主植物。其中,在棉花寄主上扶桑绵粉蚧雌成虫存活30.45±0.57 d,雄成虫存活2.41±0.19 d,每雌平均产卵量为463.73±7.91 粒,这些参数均较Fand 等(2010)在23.3~30.2℃条件下以棉花为寄主的扶桑绵粉蚧雌成虫存活(42.4±5.7 d)、雄成虫(1.5±0.1 d)和每雌平均产卵量(344±82 粒)均高。推测这种生长发育参数的差异是与试验温度有关。黄芳等(2011)在恒温27℃下的研究表明,在棉花、番茄和茄子等3种经济作物上,棉花是扶桑绵粉蚧生长发育和繁殖最适宜的寄主植物,其次是茄子和番茄。本试验比较了其在5种植物上的生长发育,并发现棉花是其适宜的寄主,但番茄是其不适宜的寄主植物。

表4 五种植物上扶桑绵粉蚧的实验种群参数Table 4 Parameters of experimental population of Phenacoccus solenopsis on different plants
扶桑绵粉蚧的寄主范围很广,在不同科的植物上其生长发育及繁殖存在差异。扶桑绵粉蚧较嗜食锦葵科、菊科、茄科植物(Aheer et al.,2009;周湾等,2012),本研究表明,供试的锦葵科扶桑和棉花、菊科的豚草和三裂蟛蜞菊,均是扶桑绵粉蚧的较适宜寄主植物,无论是发育速率和生殖能力,均高于茄科的番茄。尤其是在菊科的豚草上交尾的雌虫平均产卵量最高,为511.12±10.03 粒,其次是锦葵科的扶桑上平均产卵量为495.98±12.54 粒,比在茄科的番茄上的平均产卵量342.63±7.09 粒高1.49倍和1.45倍。在豚草上,扶桑绵粉蚧净增殖率R0为274.33,内禀增长率rm(0.18)与周限增长率λ(1.20)最大,平均世代周期T、种群加倍时间t 最短(31.44、3.88)。研究结果表明,豚草是供试植物中最适合扶桑绵粉蚧生长发育和繁殖的寄主植物,番茄为最不适合的寄主植物。
此外,在同科的植物上扶桑绵粉蚧生长发育及繁殖也存在一定差异。豚草和三裂蟛蜞菊同属于菊科,但扶桑绵粉蚧在豚草上的发育历期及寿命都显著短于三裂蟛蜞菊上的相应参数,平均产卵量则显著高于三裂蟛蜞菊上的。在锦葵科的扶桑和棉花,扶桑绵粉蚧雌虫和雄虫各个龄期的发育时间、寿命均无显著差异,但扶桑上每雌平均产卵量为495.98±12.54 粒,棉花上每雌平均产卵量为463.73±7.91 粒,两者差异显著。
汉中职业技术学院肖宁的《学前教育专业校企合作共建机制的探索与实践》一文中提出“共建人才培养方案、共建实践教学基地、共建课程、共建课堂”四建的全新观点。在这当中校企、校园要共同参与人才培养的测评及教学管理的工作,共同制定考核标准与行业准入标准,设定相似的教学环境与工作环境,统一教学内容和工作任务,了解教师与岗位工作者的要求,切实落实完善“校企共育,全程实践”的人才培养模式。
扶桑绵粉蚧寄主范围广为其发生扩散及作物闲耕期存活提供了充足的生存条件。据报道,在印度的棉区银胶菊、苍耳等杂草是扶桑绵粉蚧重要的越冬中间寄主,为次年该粉蚧在棉田发生提供了充足的虫源(Saini et al.,2009)。茄科的洋金花 Datura metal,菊科的苍耳,藜科的藜Chenopodium album,蒺藜科的蒺藜Tribulus terrestris等在夏季该害虫扩散过程中起到辅助作用,而苋科的鸡冠花 Celosia argentia,菊科的金盏花Calendula officinalis,茄科的夜香树 Cestrum nocturnum 等则是其越冬寄主(Arif et al.,2009)。这些研究报道均表明杂草是扶桑绵粉蚧传播扩散过程中的媒介,并在不良条件下(越冬或作物闲耕期)充当中间寄主。豚草和三裂蟛蜞菊均为入侵的菊科杂草,而且是广布种,已广泛分布在华东、华南地区。其中豚草已在江西、福建、湖南、广西、广东等20 多个省市分布,在广东省的韶关、清远、梅州、汕头和广州等地区分布(齐国君等,2011)。试验表明,这两种杂草是扶桑绵粉蚧较好的寄主植物,其中豚草对扶桑绵粉蚧的适生性更强,因此,这些植物可为扶桑绵粉蚧远距离扩散或作物闲耕期的存活提供充足的寄主植物。
本研究从扶桑绵粉蚧在五种植物上的适生性开展了试验,明确了植物对该虫生长发育和繁殖的影响,分析了植物对其扩散传播及暴发成灾的作用。扶桑绵粉蚧对寄主植物的选择性及其内在机制尚需进一步研究。
References)
Aheer GM,Shah Z,Saeed M,2009.Seasonal history and biology of cotton mealybug,Phenacoccus solenopsis Tinsley.Journal of Agricultural Research,47(4):423-431.
Akintola AJ,Ande AT,2008.First record of Phenacoccus solenopsis Tinsley(Hemiptera:Pseudococcidae)on Hibiscus rosa-sinensis in Nigeria.Agricultural Journal(Pakistan),3(1):1-3.
Arif MJ,Rafiq M,Ghaffar A,2009.Host plants of cotton mealybug(Phenacoccus solenopsis):a new menace to cotton agroecosystem of Punjab.International Journal of Agriculture & Biology,11(2):163-167.
Cockerell TDA,1902.Two new mealy-bugs from New Mexico.The Canadian Entomologist,34(2):315-316.
Culik MP,Gullan PJ,2005.A new pest of tomato and other records of mealybugs(Hemiptera:Pseudococcidae)from Espirito Santo,Brazil.Zootaxa,964:1-8.
Dutt U,2007.Mealy bug infestation in Punjab:Bt.Cotton falls flat.Online article at http://www.Countercurrents.org/dutt210807.htm.August.
Fand BB,Gautam RD,Chander S,Suroshe SS,2010.Life table analysis of the mealybug,Phenacoccus solenopsis Tinsley(Hemiptera:Pseudococcidae)under laboratory conditions.Journal of Entomological Research,34(2):175-179.
Granara de Willink MC,2003.New records and host plants of Phenacoccus solenopsis Tinsley for Argentina(Hemiptera:Pseudococcidae).Revista de la Sociedad Entomologica Argentina,62(3/4):80-82.
Huang F,Zhang PJ,Zhang JM,Zhu YY,Lu YB,Zhang ZJ,2011.Effects of three host plants on the development and reproduction of Phenacoccus solenopsis.Plant Protection,37(4):58-62.[黄芳,张蓬军,章金明,朱艺勇,吕要斌,张治军,2011.三种寄主植物对扶桑绵粉蚧发育和繁殖的影响.植物保护,37(4):58-62]
Hodgson C,Abbas G,Arif MJ,Saeed S,Karar H,2008.Phenacoccus solenopsis Tinsley(Sternorrhyncha:Coccoidea:Pseudococcidae),an invasive mealybug damaging cotton in Pakistan and India,with a discussion on seasonal morphological variation.Zootaxa,1913:1-35.
Larrain SP,2002.Insect and mite pest incidence on sweet pepinos(Solanum muricatum Ait.)cultivated in the IV Region,Chile.Agricultura Tecnica,62(1):15-26.
Ma J,Hu XN,Peng ZhQ,Liu HJ,Liang F,Lu YY,2011.The potential geographical distribution of Phenacoccus solenopsis Tinsley based on the CLIMEX in China.Plant Quarantine,25(1):5-8.[马骏,胡学难,彭正强,刘海军,梁帆,陆永跃,2011.基于CLIMEX 模型的扶桑绵粉蚧在中国潜在地理分布预测.植物检疫,25(1):5-8]
Nagrare VS,Kranthi S,Biradar VK,Zade NN,Sangode V,Kakde G,Shukla RM,Shivare D,Khadi BM,Kranthi KR,2009.Widespread infestation of the exotic mealybug species,Phenacoccus solenopsis(Tinsley)(Hemiptera:Pseudococcidae),on cotton in India.Bulletin of Entomological Research,99(5):537-541.
Qi GJ,Huang DC,Gao Y,Wang L,Shao XY,Sun JX,Lu LH,2011.Occurrence and distribution of Ambrosia antemisiifolia and its two natural enemies in Guangdong Province.Chinese Journal of Applied Entomology,48(1):197-201.[齐国君,黄德超,高燕,王琳,邵晓迎,孙嘉祥,吕利华,2011.广东省豚草及两种天敌昆虫的发生与分布.应用昆虫学报,48(1):197-201]
Saini RK,Ram P,Sharma SS,Rohilla HR,2009.Mealybug(Phenacoccus solenopsis Tinsley) and its survival in cotton ecosystem in Haryana.Proc.National Symposium on Bt-cotton:Opportunities and Prospects.Central Institute for Cotton Research,Nagpur.,102-103.
Tang QY,Feng MG,2007.DPS Data Processing System-Experimental Design,Statistical Analysis and Data Mining.Beijing :Science Press,43-53.[唐启义,冯明光,2007.实用统计分析及其DPS 数据处理系统.北京:科学出版社.43-53]
Tinsley JD,1898.An ants,-nest coccid from New Mexico.The Canadian Entomologist,30(2):47-48.
Vennila S,Deshmukh AJ,Pinjarkar D,Agarwal D,Ramamurthy W,Joshi S,Kranthi KR,Bambawale OM,2010.Biology of the mealybug,Phenacoccus solenopsis on cotton in the laboratory.Journal of Insect Science,10:115.
Wang YP,Wu SA,Zhang RZ,2009.Pest risk analysis of a new invasive pest,Phenacoccus solenopsis,to China.Chinese Bulletin of Entomology,46(1):101-106.[王艳平,武三安,张润志,2009.入侵害虫扶桑绵粉蚧在中国的风险分析.昆虫知识,46(1):101-106]
Wang YY,Xu ZH,Zhang LL,Shen LZ,Fu SL,2012.Developmental duration and life table of the laboratory population of Phenacoccus solenopsis Tinsley(Hemiptera:Pseudococcidae)fed on pumpkin at different temperatures.Acta Entomologica Sinica,55(1):77-83.[王莹莹,徐志宏,张莉丽,沈励泽,傅思丽,2012.南瓜寄主上扶桑绵粉蚧不同温度下的发育历期和实验种群生命表.昆虫学报,55(1):77-83]
Williams DJ,Granara de Willink MC,1992.Mealybugs of central and south America.CAB International,635.
Wu SA,Zhang RZ,2009.A new invasive pest,Phenacoccus solenopsis,threatening seriously to cotton production.Chinese Bulletin of Entomology,46(1):159-162.[武三安,张润志,2009.威胁棉花生产的外来入侵新害虫—扶桑绵粉蚧.昆虫知识,46(1):159-162]
Xu RM,1987.The Ecology of Insect Population.Beijing:Normal University Press.97-107.[徐汝梅,1987.昆虫种群生态学.北京:北京师范大学出版社.97-107]
Xu W,Fu HB,Long QH,Jiang LR,Zhao G,Zou XW,Han YC,2009.The pest,Phenacoccus solenopsis,was discovered in Hainan Province.Plant Quarantine,23(5):33.[徐卫,付海滨,龙琼华,蒋林蓉,赵刚,邹细万,韩玉春,2009.海南省发现有害生物-扶桑绵粉蚧.植物检疫,23(5):33]
Zhou W,Wang DZ,Qiu ZL,Zhou P,Liu WT,Ge X,Hu SL,Fang ZX,2012.Host plants of Phenacoccus solenopsis in Zhejiang Province and its occurrence characters.Plant Protection,38(2):152-155.[周湾,王道泽,仇智灵,周平,刘伟塘,葛翔,胡树良,方镇许,2012.扶桑绵粉蚧在浙江的寄主植物与发生特点.植物保护,38(2):152-155]
