模拟氮沉降对两种草地植物氮同化物积累的影响
2013-03-14徐瑞阳王晓红
徐瑞阳,白 龙,王晓红,刘 英
(沈阳农业大学园艺学院,辽宁 沈阳 110866)
随着氮沉降的日益加剧,氮沉降对陆地植物的影响越来越受到生态学者的关注[1-2]。氮是植物生长所需的三大养分元素之一,也是植物生长的主要限制因素之一。然而植物对氮的需求量与同化能力是有限的。当氮过量时,植物吸收的氮素量增加,氮代谢酶活性及氮同化物积累过程发生改变,最终影响植物生长[3-4]。国内外,氮沉降与森林植物的研究有了长足的发展,积累了很多研究经验[1-7],但氮沉降与草地植物的研究却鲜见报道。目前的研究主要集中在氮沉降与草地土壤呼吸、光合能力、群落结构等的关系[1,8-10]。对科尔沁沙质草地的研究表明,施氮会改变群落优势种以及物种科属组成结构,促进生产力增加[10]。沈健林等[11]研究葎草(Humulusscandens)的叶氮含量与大气活性氮的关系时证实,叶氮对大气活性氮浓度有较好的指示作用。万宏伟等[8]对羊草(Leymuschinensis)群落的研究表明,氮输入改变不同种群叶氮和叶绿素含量,进而导致群落结构的演变。蒋琦清等[12]研究表明,4 g N·m-2·a-1范围内的氮添加可显著促进禾本科和豆科草种的生物量,但不同功能类群的植株含氮及植株吸收氮的总量却无明显规律。在大气有效性氮素增加的背景下,研究植物的氮代谢功能及其氮沉降的响应,有利于掌握全球氮循环的演变过程。栽培草地在人工施氮和大气氮沉降的共同作用下往往处于氮过量的环境中。因此,本研究选取广泛种植的白三叶(Trifoliumrepens)和无芒雀麦(Bromusinermis)为研究对象,探究氮沉降背景下豆科和禾本科优质牧草氮同化物的体内积累特征及变化趋势,并探讨草本植物的氮积累指示剂,以期为研究氮沉降对草地植物氮代谢功能的影响提供科学依据。
1 材料与方法
1.1试验地概况 本试验在沈阳农业大学“百草园”教学科研基地进行。气候属于温带季风性气候,年降水量平均为677 mm,全年平均气温8.3 ℃,≥10 ℃的活动积温历年平均为3 450 ℃·d,日照时数年总量平均为2 473 h[13]。土壤为棕壤土,全氮、全磷和全钾含量分别为1.02、0.38和29.4 g·kg-1[14]。
1.2试验材料与试验设计 试验材料为沈阳农业大学“百草园”教学科研基地内种植的白三叶和无芒雀麦。本试验采用随机区组设计,基于目前我国氮沉降量逐年增加的趋势[15-17],设4个氮沉降梯度,即低氮N1(6 g N·m-2·a-1),中氮N2(12 g N·m-2·a-1)、高氮N3(24 g N·m-2·a-1)和对照N0(0 g N·m-2·a-1)。小区面积4 m × 3 m,重复3次,共12个小区。尿素为氮处理剂,整个生长季节处理10次。将年施用尿素量平均分成10等分,每次处理时将所需尿素溶解到定量的水中,用喷雾器均匀喷施于相应小区。对照只喷施等量的水。于2011年6月1日至10月15日期间,每隔15 d处理1次,共处理10次。处理7 d后取样,本文中的结果为最后一次处理后取样测得。
1.3测定项目及方法 叶绿素含量采用80%丙酮浸提法测定[18],叶氮含量采用半自动凯氏定氮仪法测定[19],可溶性蛋白含量采用考马斯亮蓝G-250染色法测定[18],精氨酸含量采用甲萘酚次氯酸钠分光光度法测定[20],脯氨酸含量采用茚三酮比色法测定[18]。
1.4统计软件 采用统计软件SPSS 12.0.1 for Windows 中单因素方差分析(One way ANOVA)和Duncan多重比较(Duncan’s multiple range test)检验不同处理间的差异。绘图制作工具Microsoft Excel。
2 结果与分析
2.1氮沉降对叶绿素含量的影响 白三叶叶绿素含量各处理极显著高于对照(P<0.01)。与对照比较,白三叶叶绿素含量增加7.7%~14.0%,中氮和高氮处理间无显著差异。而无芒雀麦的叶绿素含量随氮浓度增加而增加,各处理分别比对照增加了152.2%、225.4%和284.8%,不同处理间差异极显著(P<0.01)。可见两种牧草的叶绿素含量随氮沉降增加而增加,但两者在高氮处理下表现不同,且无芒雀麦叶绿素增量明显高于白三叶(表1)。
2.2氮沉降对叶氮含量的影响 中氮和高氮处理极显著提高了白三叶叶氮含量(P<0.01),但是这两处理间无显著差异。而无芒雀麦各处理叶氮含量均极显著高于对照(P<0.01),且N3>N2>N1>N0。无芒雀麦叶氮的积累对模拟氮沉降的响应相对于白三叶更加明显,且无芒雀麦对低浓度氮处理响应极显著(表1)。
2.3氮沉降对可溶性蛋白含量的影响 白三叶各处理可溶性蛋白含量在42.37~49.11 mg·g-1范围内,各处理均极显著高于对照(P<0.01),峰值出现在12 g N·m-2·a-1,呈明显的“单峰曲线”特征。无芒雀麦各处理可溶性蛋白质含量在33.05~43.75 mg·g-1范围内,各处理均极显著高于对照(P<0.01)。白三叶可溶性蛋白含量在高氮处理下出现极显著下降(P<0.01),而无芒雀麦可溶性蛋白含量中氮和高氮处理间无显著差异(P>0.05)(表1)。
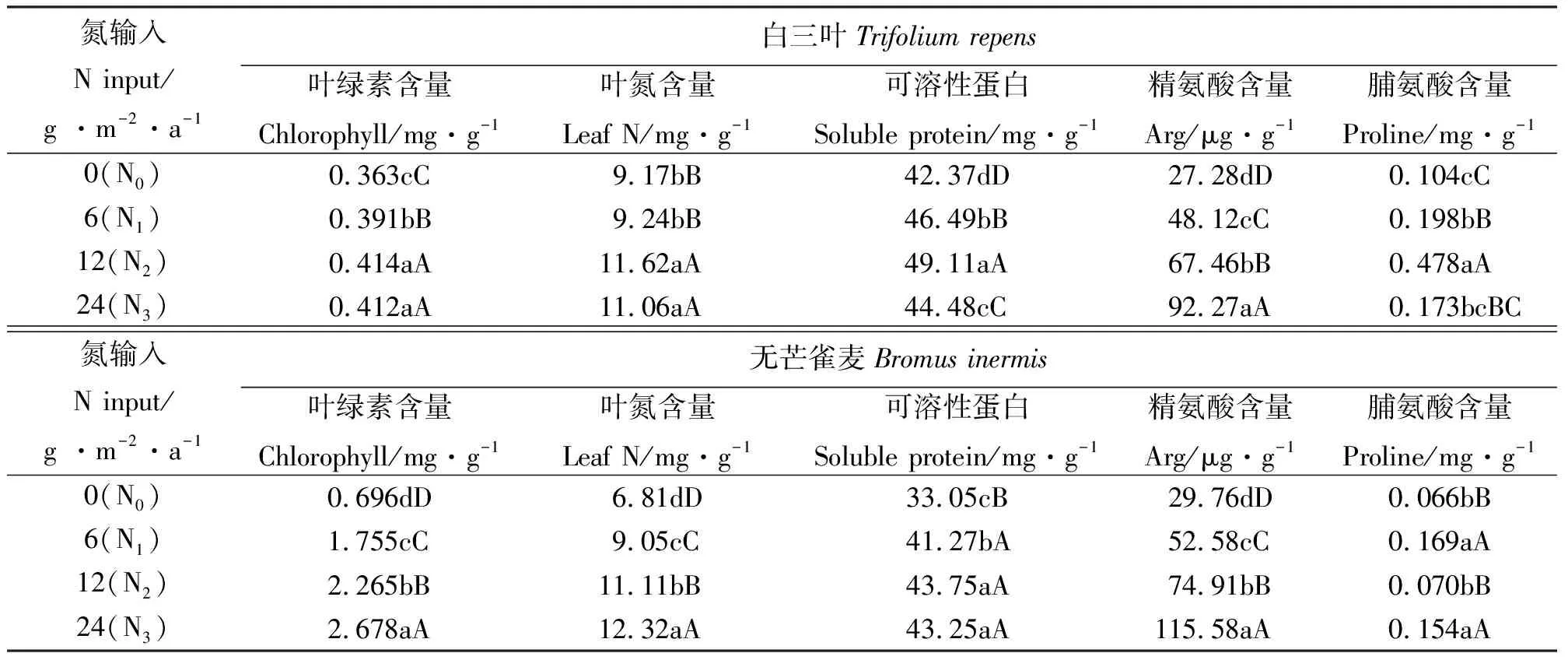
表1 氮沉降对白三叶和无芒雀麦氮同化物的影响Table 1 Effects of N input on nitrogen assimilation of Bromus inermis and Trifolium repens
2.4氮沉降对精氨酸和脯氨酸含量的影响 白三叶和无芒雀麦各处理间精氨酸含量均极显著高于对照(P<0.01),且N3>N2>N1>N0。白三叶在低、中和高氮处理中精氨酸含量分别为对照的1.76倍、2.47倍和3.38倍,而无芒雀麦的分别为对照的1.77倍、2.52倍和3.88倍,比白三叶增量更为明显(表1)。
低、中氮处理中白三叶脯氨酸含量分别比对照增加了90.4%、359.6%(P<0.01),呈现以中氮处理为峰值的单峰曲线。而无芒雀麦的对照与低氮、高氮处理差异极显著(P<0.01),与中氮处理差异不显著(P>0.05)(表1)。
相关性分析表明,氮沉降浓度与白三叶精氨酸含量间存在极显著的曲线正相关关系,而与无芒雀麦的精氨酸含量间极显著的线性正相关关系(图1)。
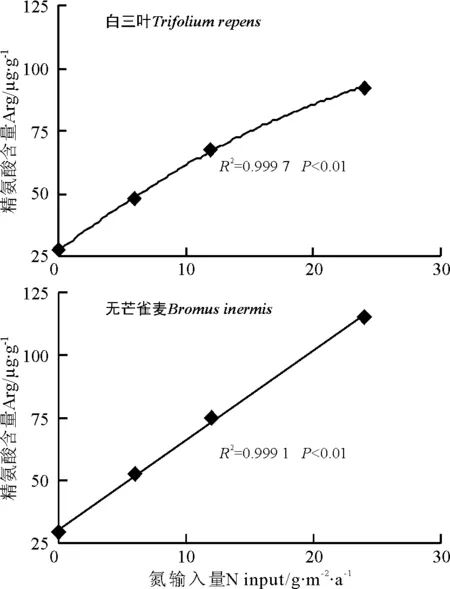
图1 氮输入量与精氨酸含量的关系Fig.1 Relationship between N input and Arg contents
3 讨论
3.1两种牧草叶绿素含量对氮沉降的响应 绿色植物的叶绿素合成受氮素和矿质元素的影响,其中氮和镁是构成叶绿素分子的重要元素[21]。研究表明,适当的氮沉降可使植物叶氮和叶绿素含量显著增加,但氮过量后反而下降[4,22-23]。本试验中,禾本科牧草的叶绿素含量随氮沉降增加而显著增加,而豆科牧草却在12 g N·m-2·a-1后不再增加。无芒雀麦的叶绿素含量最高增幅为3.8倍,而白三叶只增加1.1倍。表明在可利用氮充足的条件下,禾本科牧草叶绿素积累效应比豆科牧草更加明显。而豆科牧草的中氮和高氮处理无显著差异,可能是由于豆科牧草在大量吸收氮后,其体内增加的NH4+与其他阳离子养分进行交换,使阳离子养分从叶片中淋洗出来,造成植物体内其他养分出现“稀释效应”,导致叶片养分失衡[24-25],而养分失衡会导致叶绿素合成受阻,使豆科牧草叶绿素不再随氮处理浓度的增加而增加。在高氮处理时禾本科牧草叶绿素极显著增加,说明禾本科牧草没有发生“稀释效应”,再次证明这种“稀释效应”强度因植物种类而异[26-27]。
3.2两种牧草叶氮含量对氮沉降的响应 叶氮是植物氮同化的主要产物,其积累量与硝酸还原酶(NR)、谷氨酰胺合成酶(Gs)和谷氨酸脱氢酶(GDH)等氮同化酶的活性密切相关。通常情况下,植物叶氮含量随外界环境中有效性氮的增加而增加[28]。适当提高氮素水平可提高硝酸还原酶、谷氨酰胺合成酶和谷氨酸脱氢酶等氮同化酶的活性[29-30],而这些酶活性越高,植物根系吸收氮素的能力越强[28]。但是,当氮素水平过高时谷氨酰胺合成酶和谷氨酸脱氢酶的活性反而会下降[31]。本试验中,无芒雀麦的叶氮含量随氮沉降增加比白三叶更明显。这可能是本试验中4种氮素水平下两种牧草氮同化酶活性不同的缘故,亦表明氮沉降对禾本科牧草氮同化酶活性的作用高于豆科牧草。研究表明,土壤中过高的硝酸盐含量对豆科植物固氮的抑制作用较大[32]。本试验中豆科牧草在氮处理浓度达到12 g N·m-2·a-1后叶氮含量不再增加,可能是高氮处理使土壤中硝酸盐含量达到了抑制豆科植物固氮的水平。另有研究表明,叶氮含量增加对植物未必都有利[6,9]。氮沉降增加对两种牧草叶氮含量的影响,以及叶氮含量变化对牧草生长的影响,有待进一步研究。
3.3两种牧草可溶性蛋白质对氮沉降的响应 植物体内的可溶性蛋白质大多数是参与各种代谢的酶类,在氮素代谢中起着代谢库的作用,所以其含量是植物体总代谢能力的一个重要指标[33]。大量研究表明,氮沉降会增加植物叶片中可溶性蛋白的含量[4,22,34],且叶片中可溶性蛋白质含量与硝酸还原酶及谷氨酰胺合成酶的活性呈极显著相关[22]。通过氮添加来提高土壤中可利用氮素含量可以增加植物体内氮同化关键酶的活性和含量,从而促进氮同化物的积累和转运[22,29]。本研究中,豆科牧草可溶性蛋白质含量在氮沉降超过一定浓度后显著下降。这可能是由于过高的氮素水平会抑制其代谢酶的合成,并且抑制氮同化酶的活性。但无芒雀麦并未出现此类现象,并且其可溶性蛋白含量增幅比白三叶高近两倍。这意味着一定范围内的氮沉降显著促进两种牧草可溶性蛋白的合成和积累,且氮沉降对禾本科牧草的氮同化物积累的促进作用高于豆科牧草。
3.4两种牧草氮沉降指示剂 过量的氮在植物体内主要以游离氨基酸的形式积累似乎是一种有效的贮存多余氮的途径[4,6-7]。而精氨酸是唯一含有4个氮原子的氨基酸,因而成为更有效的氮贮存形式[5-6]。氮沉降增加对精氨酸的影响在本研究中也得到进一步的证实。两种牧草的精氨酸含量皆随氮处理浓度的增加呈现极显著的上升趋势(P<0.01),其中高氮处理比对照分别增加了2 382.3%和2 883.7%。氮沉降浓度与白三叶精氨酸含量间存在极显著的曲线正相关关系,而与无芒雀麦的精氨酸含量间存在极显著的线性正相关关系。脯氨酸含量未出现明显的变化规律。表明,两种牧草中,精氨酸为贮存多余氮的主要方式,可作为草本植物对氮沉降增加的敏感指标。
综上所述,氮沉降对两种牧草的叶绿素、叶氮、可溶性蛋白、精氨酸等氮同化物积累有一定的影响,并且两种牧草的响应机制有差异。需要进行进一步的长期监测。
[1] 张燕,崔学民,樊明寿.大气氮沉降及其对草地生物多样性的影响[J].草业科学,2007,24(7):12-17.
[2] 张维娜,廖周瑜.氮沉降增加对森林植物影响的研究进展[J].环境科学导刊,2009,28(3):21-24.
[3] Kontunen-Soppela S,Taulavuori K,Taulavuori E,etal.Soluble proteins and dehydrins in nitrogen-fertilized Scots pine seedlings during deacclimation and the onset of growth[J].Physiologia Plantarum,2000,109(4):404-409.
[4] Warren C R,Dreyer E,Adams M A.Photosynthesis-rubisco relationships in foliage ofPinussylvestrisin response to nitrogen supply and the proposed role of Rubisco and amino acids as nitrogen stores[J].Trees,2003,17(4):359-366.
[5] 鲁显楷,莫江明,彭少麟,等.鼎湖山季风常绿阔叶林林下层3种优势树种游离氨基酸和蛋白质对模拟氮沉降的响应[J].生态学报,2006,26(3):743-753.
[6] Calanni J,Berg E,Wood M,etal.Atmospheric nitrogen deposition at a conifer forest:Response of free amino acids in Engclmann spruce needles[J].Environmental Pollution,1999,l05(1):79-89.
[7] Brown K R,Thompson W A,Camm E J,etal.Effects of N addition rates on the productivity ofPiceasitchensis,Thujaplicata,andTsugaheterophyllaseedlings II.Photosynthesis,13C discrimination and N partitioning in foliage[J].Trees,1996,10(3):198-205.
[8] 万宏伟,杨阳,白世勤,等.羊草草原群落6种植物叶片功能特性对氮素添加的响应[J].植物生态学报,2008,32(3):611-621.
[9] 朱天鸿,程淑兰,方华军,等.青藏高原高寒草甸土壤CO2排放对模拟氮沉降的早期响应[J].生态学报,2011,31(10):2687-2696.
[10] 李禄军,于占源,曾德慧,等.施肥对科尔沁沙质草地群落物种组成和多样性的影响[J].草业学报,2010,19(2):109-115.
[11] 沈健林,黄明丽,唐傲寒,等.葎草(Humulusscandens)叶总氮和δ15N与大气活性氮气体的关系[J].中国农业大学学报,2010,15(1):84-88.
[12] 蒋琦清,唐建军,陈欣,等.模拟氮沉降对杂草生长和氮吸收的影响[J].应用生态学报,2005,16(5):951-955.
[13] 刁军,董晓明,刘凤芝,等.近49年沈阳东陵区光温水资源特征与趋势变化.安徽农业科学,2010,38(34):19634-19635,19698.
[14] 赵野,苏芳莉,崔彬,等.土壤前期含水量对棕壤土坡耕地养分流失的影响.水土保持学报,2011,25(1):25-29.
[15] 谢迎新,张淑利,冯伟,等.大气氮素沉降研究进展[J].中国生态农业学报,2010,18(4):897-904.
[16] 孙志高,刘景全,王金达.三江平原典型湿地系统湿沉降中氮浓度及沉降量初步研究[J].农业系统科学与综合研究,2007,23(1):114-119,123.
[17] 宇万太,马强,张璐,等.下辽河平原降雨中氮素的动态变化[J].生态学杂志,2008,27(1):33-37.
[18] 李玲.植物生理学模块实验指导[M].北京:科学出版社,2010:37-86.
[19] 张丽英.饲料分析及饲料质量检测技术[M].北京:中国农业出版社,2004:49-55.
[20] 胡桂娟,刘寄明,刘嘉芬.化学法测定精氨酸总量[J].落叶果树,1995(1):22.
[21] 萧浪涛,王三根.植物生理学[M].北京:中国农业出版社,2004:126-136.
[22] 李晶.密度与氮素水平对小黑麦氮代谢及产量品质的影响[D].哈尔滨:东北农业大学,2009:91-94.
[23] 解宝,陈果,杜金鸿,等.氮与pH互作对菊苣4项生理指标的影响[J].草业科学,2012,29(4):571-576.
[24] Van der Eerden L J,Lekkerkerk L J A,Smeulders S M.Effects of atmospheric ammonis and ammonium phate in forest canopy throughfall[J].Nature,1982,299:548-550.
[25] Hüttl R F.Nutrient supply and fertilizer experiments in view of N saturation[J].Plant Soil,1990,128(1):45-58.
[26] 李德军,莫江明,彭少麟,等.南亚热带森林两种优势树种幼苗的元素含量对模拟氮沉降增加的响应[J].生态学报,2005,25(9):2165-2172.
[27] Nakaji T,Fukami M,Dokiya Y,etal.Effects of high nitrogen load on growth,photosynthesis and nutritrient status ofCryptomeriajaponicaandPinusdensiflraseedlings[J].Trees,2001,15(8):453-461.
[28] 杨红丽,陈功,吴建付.施氮水平对多花黑麦草植株氮含量及反射光谱特征的影响[J].草业学报,2011,20(3):239-244.
[29] Cren M,Hirel B.Glulamine synthetase in higher plant:Regulation of gene and protein expression from the organ to the cell[J].Plant and Cell Physiology,1999(40):1187-1193.
[30] 王平.不同氮效率类型小麦氮代谢差异及其机理分析[D].泰安:山东农业大学,2011:100-101.
[31] 张智猛,万书波,宁堂原,等.氮素水平对花生氮素代谢及相关酶活性的影响[J].植物生态学报,2008,32(6):1407-1416.
[32] 肖焱波,李隆,张福锁.豆科——禾本科间作系统中氮营养研究进展[J].中国农业科技导报,2003,5(6):44-49.
[33] 于亚军,李军,贾志宽,等.旱作农田水肥耦合研究进展[J].干旱地区农业研究,2005,23(3):220-224.
[34] 丁雪梅,苑兆和,冯立娟,等.不同氮素水平对大丽花形态指标及生理特性的影响[J].草业科学,2012, 29(4):592-598.
