4个核桃品种光合特性的日变化
2013-03-14宗建伟杨雨华杨风岭梁亚红刘杜玲朱海兰
宗建伟,杨雨华,杨风岭,梁亚红,刘杜玲,朱海兰
(1.平顶山学院资源与环境科学学院 低山丘陵生态修复重点实验室,河南 平顶山 467000; 2.西北农林科技大学林学院,陕西 杨凌 712100)
核桃(Juglansregia)为胡桃科核桃属,它的果实不仅富含多种氨基酸,而且还含有人体必需的多种矿质元素,具有较高的营养、保健及药用价值[1]。我国是世界上核桃原产地之一,目前在我国种植的7属28种胡桃科植物中,有4个种已经确定为是原产于我国的核桃品种。由于核桃具有很高的经济价值和广泛的生态适应性,核桃产业在我国得以迅猛发展。目前,对核桃的研究多集中在繁育技术、新品种引进以及病虫害防治等方面,而对光合生理性能方面的研究少有涉及。环境变化直接影响着植物的光合作用[2],而光合作用的大小则直接影响着果树干物质的积累和果实的产量、品质。目前,在研究植物光合作用过程中,光合指标已越来越受到广大研究人员的肯定和广泛应用[3-5]。本研究分析4个核桃品种的光合特性日变化,旨在通过比较4个核桃品种光合特征的差异,为优质高产提供依据。
1 材料与方法
1.1试验地概况 试验地在陕西杨凌西北农林科技大学林学院教学试验苗圃。地处108°07′ E,34°12′ N,海拔454.5 m,属暖温带半湿润大陆性季风气候,年均降水量660 mm,年均温12.9 ℃,年均积温4 811 ℃·d,无霜期220 d,年均日照时数2 163.8 h,年均辐射总量114.8 kJ·m-2。
1.2试验材料 试验材料选用4个品种1年龄早实核桃嫁接苗,4个品种分别为鲁光、扎343、强特勒、中林5号。
1.3试验设计 2011年3月上旬选择健康无病、长势一致,株高1.2 m的4个品种的嫁接核桃苗分别种植于塑料桶(高度30 cm、桶口直径25 cm)中,每个品种3桶,每桶1株。基质配比为熟土∶营养土∶沙=3∶2∶1,质量控制在18 kg左右。
1.4光合指标的测定方法 测定时间为2011年6月10日,采用由美国LI-COR公司生产的LI-6400便携式光合测定仪2 cm×3 cm透明叶室分别测定每个核桃品种的光合日变化。自08:00-18:00每间隔2 h测定1次,每个品种均选取离枝顶端向下第3~4片复叶上的第3~4片功能叶,每个品种选取5个叶片且每个叶片测定3次数据[6],然后对其求平均值。为避免仪器遮挡和光源差异较大引起的误差,测定时尽量使叶室与自然光线垂直,每测完一个叶片后对其挂牌编号,便于下次测定。测定光合参数包括:1)净光合速率(Pn,μmol·m-2·s-1)、气孔导度(Gs,mol·m-2·s-1)、蒸腾速率(Tr,mmol·m-2·s-1),2)环境因子:光合有效辐射(PAR,μmol·m-2·s-1) 、气温(Ta,℃)、大气CO2浓度(Ca,μmol·mol-1)、空气相对湿度(RH,%)。根据光合和环境参数计算水分利用效率(Water Use Efficiency,WUE)。
WUE=Pn/Tr.
1.5数据处理 绘图和数据统计分析采用Microsoft Excel 2007和SPSS 16.0软件结合进行。
2 结果与分析
2.1环境因子日变化 从08:00至18:00,光合有效辐射、气温的日变化均呈现单峰曲线,具有先升后降的趋势,光合有效辐射的峰值出现在12:00左右,达1 764.741 μmol·m-2·s-1;而气温的峰值出现在14:00左右,达41.54 ℃。空气CO2浓度在08:00-10:00出现了急剧下降的现象,而在10:00-18:00期间变化平稳;从08:00-16:00,空气相对湿度日变化出现逐渐下降的趋势(图1)。
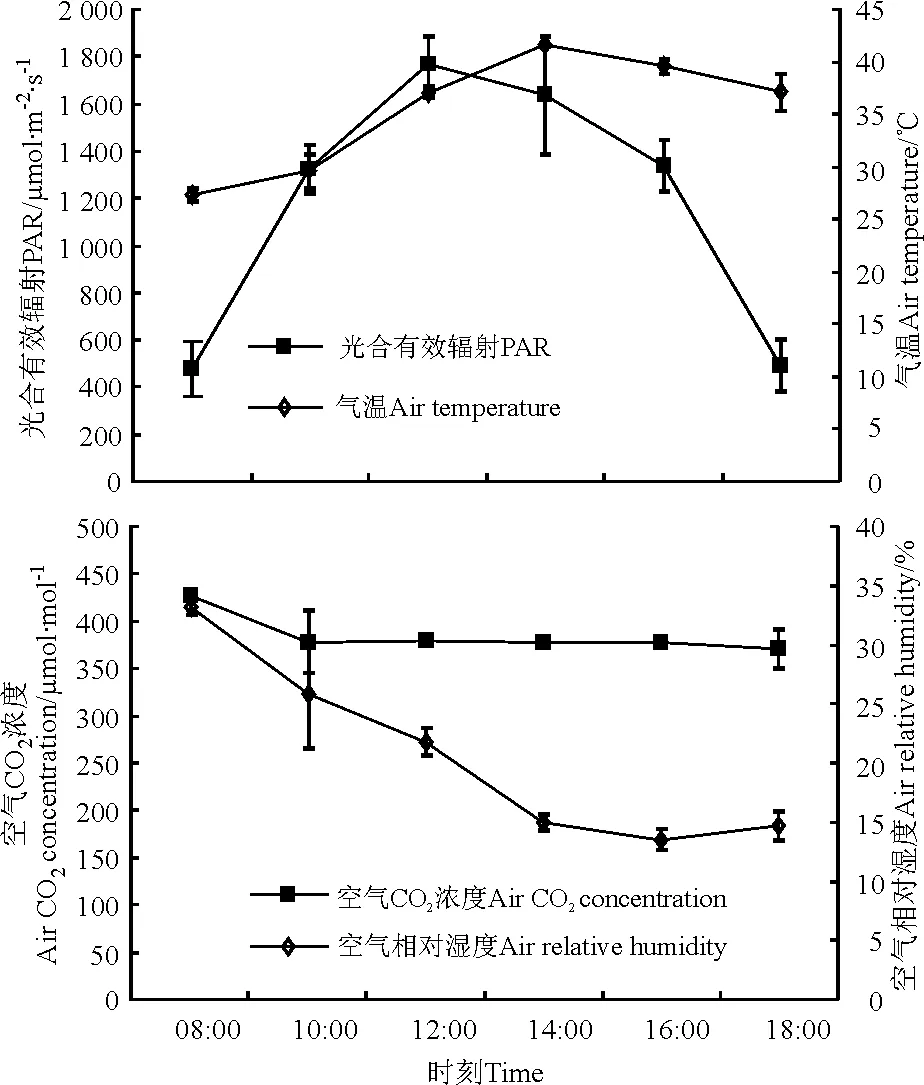
图1 光合有效辐射、空气温度、空气CO2浓度和空气相对湿度日变化规律Fig.1 Diurnal change of photosynthetically active radiation,air temperature,air CO2 concentration and air relative humidity
2.2光合特征日变化
2.2.1净光合速率日变化规律 4个不同的核桃品种净光合速率日变化均呈现双峰型曲线,且在14:00均出现了显著的“午休”现象(图2A)。不同的是,4个核桃品种净光合速率第1次峰值出现的时间并不相同,鲁光和扎343第1次峰值出现在10:00左右,其峰值分别为4.846和7.003 μmol·m-2·s-1;而强特勒和中林5号第1次峰值则出现在08:00左右,其峰值分别为6.122和5.351 μmol·m-2·s-1。4个核桃品种的第2次峰值均出现在16:00左右。此外,4个核桃品种的最大净光合速率由大到小依次排序为:扎343(7.003 μmol·m-2·s-1)>强特勒(6.122 μmol·m-2·s-1)>中林5号(5.351 μmol·m-2·s-1)>鲁光(4.846 μmol·m-2·s-1),且4个品种间不存在显著差异(P>0.05)。
2.2.2蒸腾速率日变化规律 鲁光、中林5号两个品种的蒸腾速率日变化呈现单峰曲线,峰值均出现在14:00左右,分别是2.038和1.605 mmol·m-2·s-1;而扎343、强特勒的蒸腾速率日变化呈现双峰曲线,峰值分别出现在10:00和14:00左右(图2B)。4个品种的蒸腾速率均值日变化由大到小依次排序为扎343(1.551 mmol·m-2·s-1)>强特勒(1.388 mmol·m-2·s-1)>鲁光(1.341 mmol·m-2·s-1)>中林5号(1.152 mmol·m-2·s-1)(图2B),且扎343与其余3个品种达到显著差异(P<0.05),而强特勒和鲁光之间差异不显著(P>0.05)。
2.2.3气孔导度日变化规律 鲁光、扎343的气孔导度先呈增加趋势,出现峰值之后均呈下降趋势(图2C)。然而,4个核桃品种的气孔导度峰值出现的时间点并不一致,鲁光和扎343气孔导度峰值均出现在10:00左右,分别为0.052和0.065 mol·m-2·s-1,而强特勒和中林5号峰值出现在08:00左右,其值分别为0.060和0.069 mol·m-2·s-1。其中,中林5号和强特勒、鲁光之间,扎343和鲁光之间均存在显著差异(P<0.05)。
2.2.4水分利用效率日变化规律 4个核桃品种的水分利用效率从08:00左右开始均呈现下降的趋势,到16:00左右出现了起伏,但峰值均低于08:00左右的峰值,之后又逐渐下降(图2D)。4个品种的水分利用效率均值日变化由大到小依次排序为强特勒(2.261 mmol·mol-1)>中林5号(2.190 mmol·mol-1)>扎343(1.965 mmol·mol-1)>鲁光(1.915 mmol·mol-1),且强特勒、中林5号和扎343、鲁光之间存在显著差异(P<0.05)。
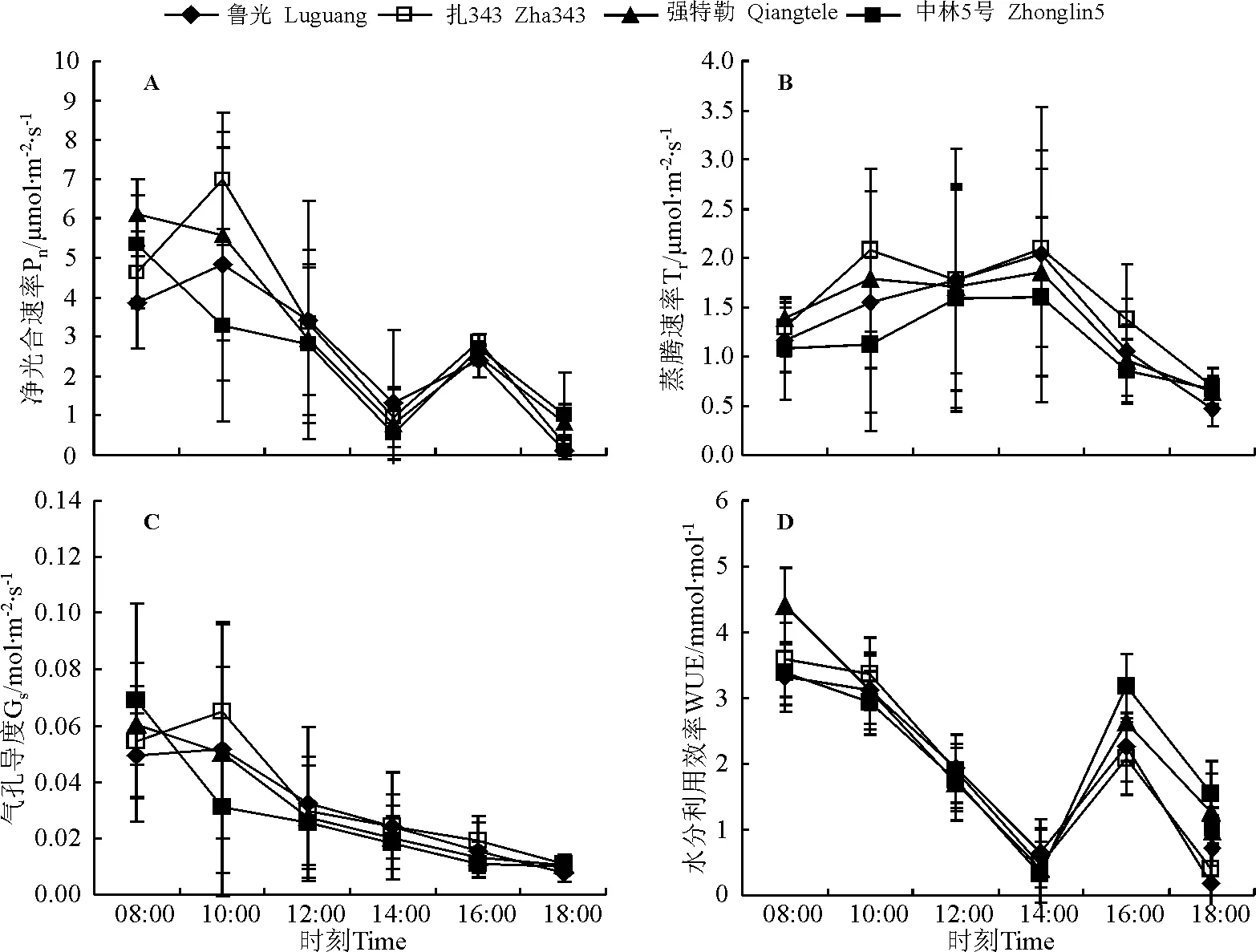
图2 4个核桃品种光合特征日变化Fig.2 Diurnal changes of photosynthetic characteristics of four Juglans regia cultivars
2.3不同核桃品种光合参数与环境指标相关性分析 4个核桃品种的净光合速率与气孔导度、水分利用效率、光合有效辐射均存在显著的正相关(P<0.05),而气温与气孔导度存在显著负相关。此外,还发现光合有效辐射与核桃品种鲁光的蒸腾速率存在显著正相关,但气温与扎343、强特勒的水分利用效率存在显著负相关,中林5号和强特勒的净光合速率与气温存在显著负相关。鲁光、扎343的气孔导度与水分利用效率存在显著正相关,而强特勒、中林5号的气孔导度与水分利用效率无相关性。
3 讨论与结论
植物叶片捕获的激发能高于植物本身的碳同化能力时,常造成过多的光能被消耗,最终引起光合速率的下降[7]。本研究发现,4个核桃品种净光合速率日变化呈现双峰曲线,且14:00左右均出现了明显的“午休”现象,由于净光合速率与光有效辐射、气孔导度存在显著的正相关(P<0.05),因此,引起中午核桃“午休”现象发生的因素光强可能并不是最主要的原因。有研究表明,环境的气温与净光合速率的相关性是一种假象[8-10],因此,气温不是直接限制净光合速率的因素之一,而且本研究也发现,鲁光和扎343两个品种的净光合速率与气温相关性不显著。
由于强特勒和中林5号净光合速率第1个峰值出现在08:00左右,而鲁光和扎343比强特勒和中林5号峰值推迟出现2 h,而且净光合速率与光合有效辐射存在显著正相关(P<0.05),且光合有效辐射峰值出现在12:00左右,这可能是强特勒和中林5号较鲁光和扎343更早地受到了光抑制,因此,强特勒和中林5号在日常管理中应注意遮阴,过大的光照强度不仅影响植物的光合能力,甚至对植物造成伤害。
在同样的环境条件下,与其他3个核桃品种相比,扎343蒸腾速率日变化呈现双峰型曲线,峰值分别出现在10:00和14:00,因此,相对于其他3个核桃品种,扎343对环境的适应具有一定的滞后性,其原因可能与不同核桃品种遗传特性差异有关。
光合有效辐射给植物碳同化提供着能量,是植物光合作用的光来源。自然条件下,植物的光合能力与光照强度之间存在正相关。有研究表明,当光合有效辐射在一天中处于较低值时,净光合速率会随着光合有效辐射的升高而增加[11-12]。本研究中,4个核桃品种净光合速率与光合有效辐射均存在显著正相关,但4个品种的净光合速率与光合有效辐射的相关系数不同。
植物叶片的光合能力不仅受自身遗传特性的影响,而且还受外部环境因子的影响[13]。本研究发现,净光合速率与气孔导度、水分利用效率、光合有效辐射均存在显著正相关(P<0.05)。因此,影响4个核桃品种的净光合速率日变化的主要因子可能是气孔导度、光合有效辐射和水分利用效率。总之,在核桃管理中不仅要注重水肥的管理,更不能忽视对核桃的整形修剪,从而保证植株采光及通风,通过综合措施减少光合午休的时间,提高光合效应。
[1] 郗荣庭,张毅萍.中国核桃[M].北京:中国林业出版社,1992:12-19.
[2] 周朝彬,宋于洋,王炳举,等.干旱胁迫对胡杨光合和叶绿素荧光参数的影响[J].西北林学院学报,2009,24(4):5-9.
[3] 王红霞,张志华,王文江,等.田间条件下核桃光合特性的研究[J].华北农学报,2007,22(2):125-128.
[4] 刘鹏,刘庆忠,赵红军,等.核桃光合作用特性的初步研究[J].落叶果树,2003,35(4):1-3.
[5] 贺奇,王贵,常月梅,等.早实核桃光合特性的初步研究[J].山西农业大学学报,2010,30(3):197-199.
[6] 刘飞虎.植物品种净光合速率测定的取样技术——以苎麻(Boehmerianivea(L.) Gaud.)为例[J].云南大学学报,2010,32(2):221-226.
[7] Mller P,Li X P,Niyogit K K.Non-photochemical quenching.A response to excess light energy[J].Plant Physiology,2001,125(4):1558-1566.
[8] 岳春雷,刘亚群.濒危植物南川升麻光合生理生态的初步研究[J].植物生态学报,1999,23(1):71-75.
[9] 张文标,金则新,柯世省,等.木荷光合特性日变化及其与环境因子相关性分析[J].广西植物,2006,26(5):492-498.
[10] 柯世省,陈模舜.紫荆光合日变化及其与环境因子的关系[J].天津师范大学学报(自然科学版),2004,24(3):30-33.
[11] Hoyaux J,Moureaux C,Tourneur D,etal.Extrapolating gross primary productivity from leaf to canopy scale in a winter wheat crop[J].Agricultural and Forest Meteorology,2008,148(4):668-679.
[12] 刘小文,王秋霞,郭美霞,等.紫茎泽兰生殖期光合特性日变化[J].草业科学,2011,28(1):105-109.
[13] 王红霞,张志华,王文江,等.河北核桃(JuglanshopeiensisHu)光合特性的研究[J].园艺学报,2005,32(3):392-396.
