新疆霍城假苇拂子茅种群构件的年龄结构
2013-03-14努尔买买提
赵 玉,刘 影,努尔买买提
(资源与生态研究所 新疆自治区普通高校天然产物化学与应用重点实验室伊犁师范学院化学与生物科学学院,新疆 伊宁 835000)
在漫长的进化过程中,植物普遍形成了自身的营养繁殖习性[1-3],尤其是多年生草本植物,必须依赖营养繁殖维系其生长及其种群或群落的稳定性[4-5]。那些通过营养繁殖可以形成独立生活子代的植物被称为无性系植物(Clonal Plant)。在温带草原地区,对于多年生草本植物而言,营养繁殖是其实现种群更新和维持群落中地位的主要方式,该过程主要通过多年生部位分蘖节和地下根茎芽的输出[6-12]作为保障。国内自20世纪80年代以来,在草原常见植物的营养繁殖特性方面陆续开展了相关研究工作,尤其是草原优势种与建群种的营养繁殖生态学特征,如羊草(Leymuschinensis)[11-13]、光稃茅香(Hierochloeglabra)[14]、碱茅(Puccineuiadistans)[15]、星星草(Puccinelliatenuiflora)[16]等。这些工作的开展,对了解多年生草本植物的营养繁殖特性和补充更新方式,以及草地资源的管理和科学利用等都具有重要理论与实践价值。
假苇拂子茅(Calamagrostispseudophragmits)为根茎型多年生禾草,可通过较强的营养繁殖特性形成大面积的单优群落型草地。在新疆伊犁河谷地区,假苇拂子茅作为该地区湿地草甸和沼泽化草甸的优势种或重要伴生种,既是重要的饲用禾草之一,也是湿地生态工程潜在的优质工程用植物。目前,有关假苇拂子茅的研究报道主要集中在其群落特征及生物量的调查[17-19]、生态位[20]、种群构件的年龄结构[21-23]、物质生产[24]和有性生殖构件特征[25-26]等方面。作为一种重要的牧草、造纸原料及优良的水土保持植物,在其种群层次上开展的营养繁殖策略及其环境适应性,以及无性系种群的生物生态学机制的研究还鲜有报道。本研究通过对霍城县大西沟河岸假苇拂子茅群落的取样调查,对3个不同生境的假苇拂子茅无性系种群开展营养繁殖特征分析,以揭示其营养繁殖与持续更新机制,进而为该地区的假苇拂子茅草地资源管理与科学利用提供理论依据。
1 研究样地与研究方法
1.1样地自然概况 试验选取的3个样地均位于新疆霍城县大西沟乡(80°75′ E,44°26′ N)沟谷地带的天然假苇拂子茅草地。该地区海拔1 000~1 150 m,年均温8.2 ℃,极端最高温40.1 ℃,极端最低温-42.6 ℃,山脚平原区年均降水量220 mm左右,无霜期165 d,年日照时数2 800 h左右[27]。该乡地形复杂,天山北麓支脉的婆罗克努山造就了山区独特的逆温带小气候特征,与山脚下平原地带相比较具有明显的冬暖夏凉的特征,年降水量500 mm左右。生境Ⅰ(H1)为大西沟河冲击河滩假苇拂子茅草地,地处逆温层带,海拔1 400 m,土壤为沙壤土,牛、羊等牲畜采食频繁,伴生种为金色狗尾草(Setariaglauca)、天蓝苜蓿(Medicagolupulina)、狗牙根(Cynodondactylon)和柽柳(Tamarixchinensis),假苇拂子茅盖度80%左右,所受干扰较轻;生境Ⅱ(H2)为山前坡地假苇拂子茅单优群落,地处平原与逆温带的过渡区域,海拔1 020 m,该生境无放牧及人为干扰,土壤潮湿、松软且土层厚,土壤为栗钙土夹杂少量沙壤土,伴生种为中国旋花(Convolvulusarvensis),假苇拂子茅盖度75%左右;生境Ⅲ(H3)为大西沟河大西沟乡段河岸季节性水淹生境,海拔820 m,土壤松软,其上层为栗钙土,中层为沙,底层为鹅卵石,松软潮湿,伴生种为芦苇(Phragmitesaustralis)、问荆(Equisetumhiemale)、红皮柳(Salixsinopurpurea)和柽柳,假苇拂子茅盖度80%左右。
1.2研究方法 2010年10月上旬,分别在3个样地内对假苇拂子茅无性系种群进行取样。样方的大小为0.25 m×0.25 m,深度为0.3 m,取样时将地上部分连同地下根茎一并挖出,重复5次,并编号分置[22,24]。保持地上与地下部构件的自然联系以及新老分蘖株的自然联系,于实验室内用清水轻轻冲洗干净,参照根茎禾草无性系种群年龄结构的研究方法[28]将样品中的分蘖株和根茎划分龄级并分别计数。各龄级分蘖株和根茎于80 ℃烘干至质量恒定,逐样方称其生物量。
1.3数据分析 数据的统计、分析和制图采用Microsoft Excel 2003及SPSS 13.0软件进行。分蘖株数及其生物量、根茎长与根茎生物量等数量指标均换算成1 m2的数据指标,并用Duncan’s法进行同一生境的不同龄级间或不同生境间相关数量指标的差异显著性检验。
2 结果与分析
2.1分蘖株的年龄结构 3个生境的假苇拂子茅种群分蘖株的年龄结构如表1和表2所示。结果表明,除生境H3外,假苇拂子茅无性系种群分蘖株均由5个龄级组成(表1),且同一生境内不同龄级间的差异显著(P<0.05)。3个生境中单位面积的总分蘖株数表现为H1>H3>H2,生境间差异显著(P<0.05);3个生境内的分蘖株均以2 a龄级占优势地位,1 a、3 a、4 a和5 a龄级所占比例依次减小。3个生境的假苇拂子茅无性系种群分蘖株在数量上均属于稳定型年龄结构。
单位面积上假苇拂子茅无性系种群的生物量表现为H2>H3>H1,不同生境间差异显著(P<0.05),且H2和H3分别是H1的2.76和1.96倍以上。在假苇拂子茅分蘖株生物量结构中,H1和H3均以1 a龄级所占比例最高,分别达到了39.47%和46.06%,而在H2中则以2 a龄级的最高。各龄分蘖株的变异系数(CV)均以5 a龄级为最高,这充分体现了3个生境的假苇拂子茅无性系种群高龄级分蘖株在不同生境中有相似的变异特点(表2)。
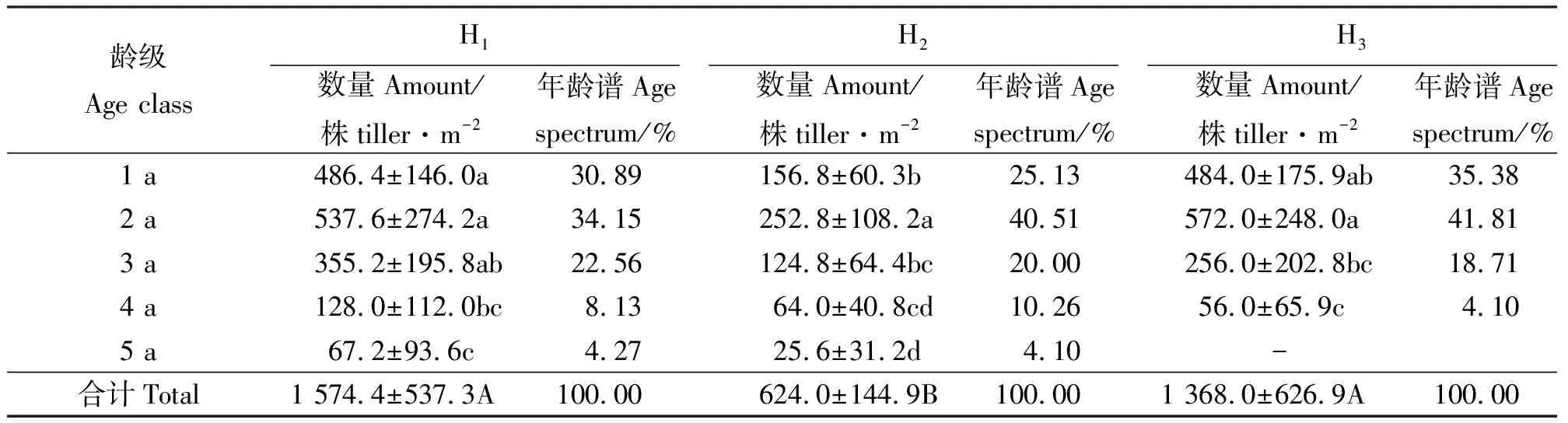
表1 不同生境假苇拂子茅种群分蘖株数量及其年龄谱
2.2不同龄级分蘖株生产力的比较 3个生境的无性系单株生产力均以1 a龄级最高,且均随着龄级的增加而依次减少,至5 a龄级最低;生境H2的1 a龄级分蘖株生产力最高,达2.181 6 g·株-1,为生产力最低的H1中1 a龄级的6.9倍,且不同龄级间差异显著(P<0.05)。从整体来看,3个生境的单株生产力水平表现为H2>H3>H1,且生境间差异显著(P<0.05),反映了单株生产力在不同植被组成间具有一定的差异(表3)。在没有放牧及人为干扰压力下,假苇拂子茅无性系种群具有旺盛的生产力水平,而频繁的放牧、水淹等干扰不利于其分蘖株的物质生产。
2.3根茎的年龄结构 3个生境中假苇拂子茅无性系种群的根茎年龄级最高均为5 a。除生境H1外,根茎长(表4)和根茎生物量(表5)均以1 a龄级所占比例最大,且二者随着龄级的增加而不断减小,龄级间差异显著(P<0.05)。3个样地单位面积的总生物量则表现为H1>H2>H3,反映出潮湿、疏松的沙质土壤有利于假苇拂子茅根茎的生长和物质储存,且生境间差异显著(P<0.05)。3个生境中的假苇拂子茅种群根茎均以1 a和2 a龄级所占比例最大,3 a和4 a次之,而以5 a最少,其根茎长度所占百分比仅为2.13%~10.80%,生物量仅为1.07%~3.13%。
2.4不同龄级根茎贮藏力的比较 通过对3个生境的假苇拂子茅无性系种群单位长度根茎的生物量即生产力的研究发现,H1中根茎的生产力以1 a龄级最高,2 a、3 a、4 a和5 a龄级的养分贮存能力不断减弱。由此反映出,在生境H1中,潮湿、疏松的沙质土壤生境,有利于假苇拂子茅无性系种群在当年形成大量的1 a根茎,并赋予其旺盛的生活力和养分贮存能力。对于生境H2和H3,则以2 a根茎的生产力最高,其次分别为1 a、3 a、4 a和5 a(表6),根茎生产力在3个生境间差异不显著(P>0.05)。
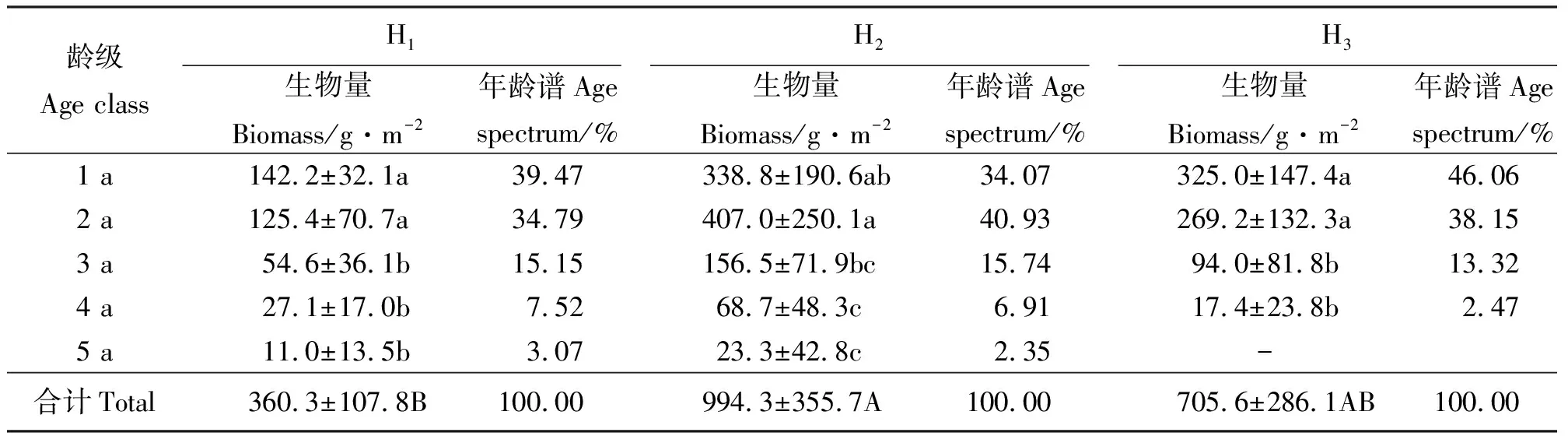
表2 不同生境假苇拂子茅种群分蘖株生物量及其年龄谱Table 2 Biomass and age spectrum on tillers of Calamagrostis pseudophragmites populations in different habitats
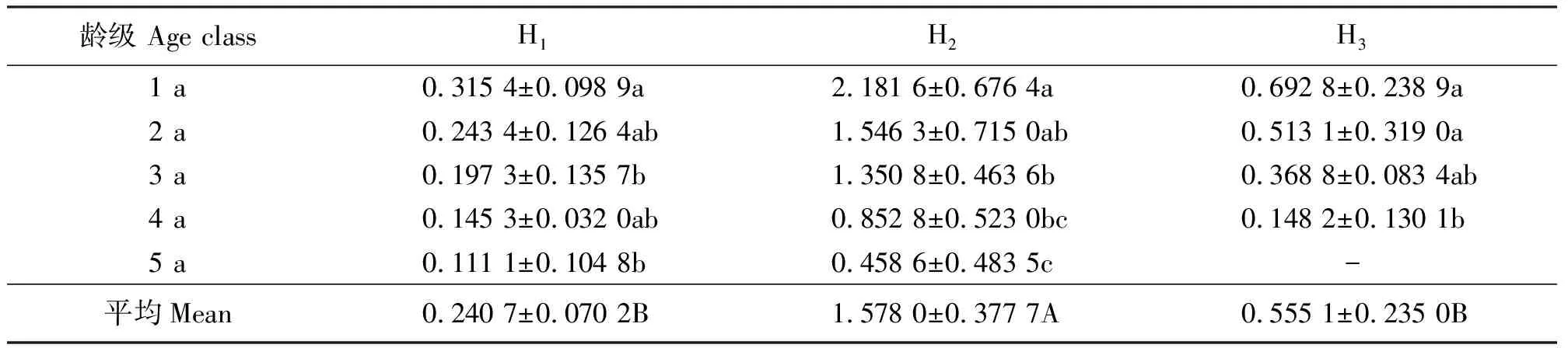
表3 不同生境假苇拂子茅无性系种群分蘖株生产力 g·株-1
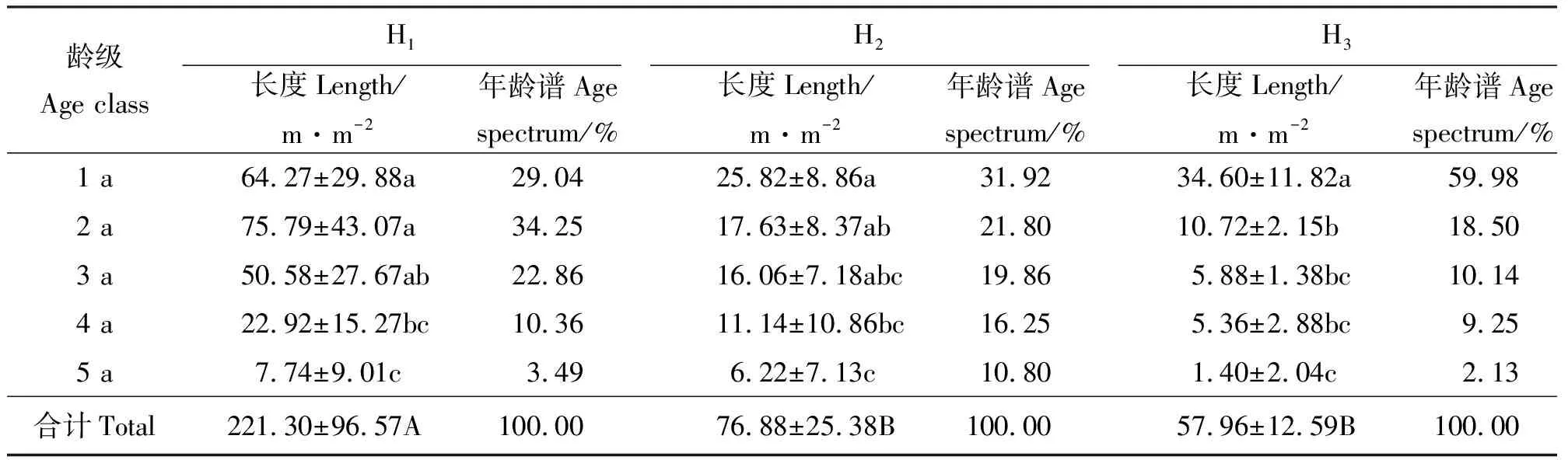
表4 不同生境假苇拂子茅无性系种群的根茎累积长度及其年龄谱Table 4 Rhizomes’ accumulated length and age spectrum of Calamagrostis pseudophragmites clonal populations in different habitats
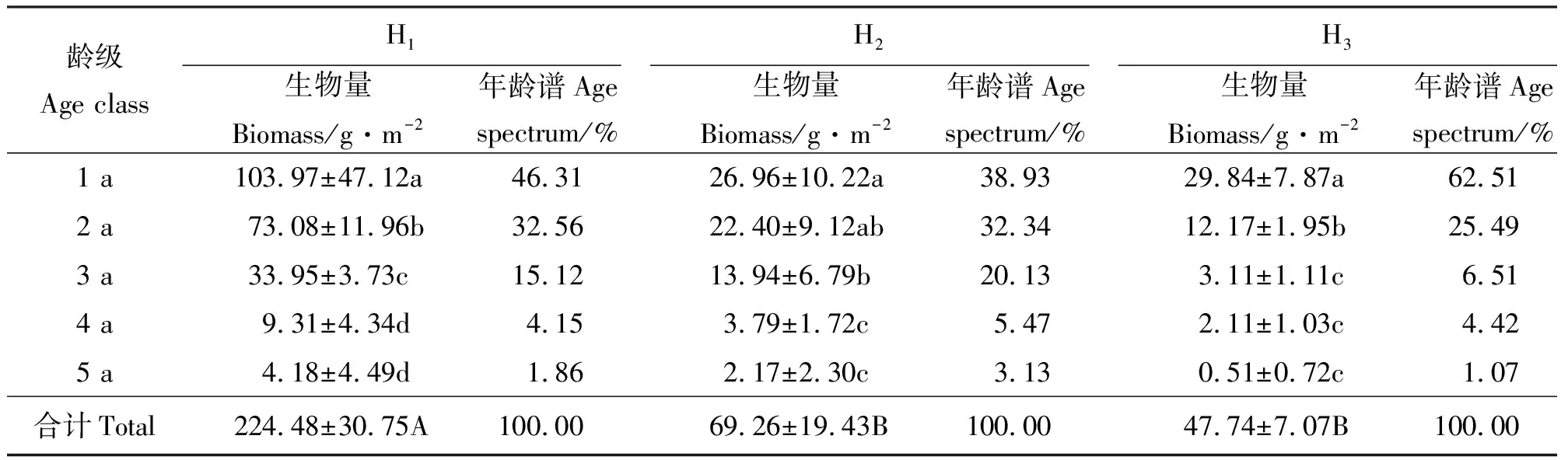
表5 不同生境假苇拂子茅无性系种群根茎的累积生物量及其年龄谱Table 5 Rhizomes’ accumulated biomass and age spectrum of Calamagrostis pseudophragmites clonal populations in different habitats
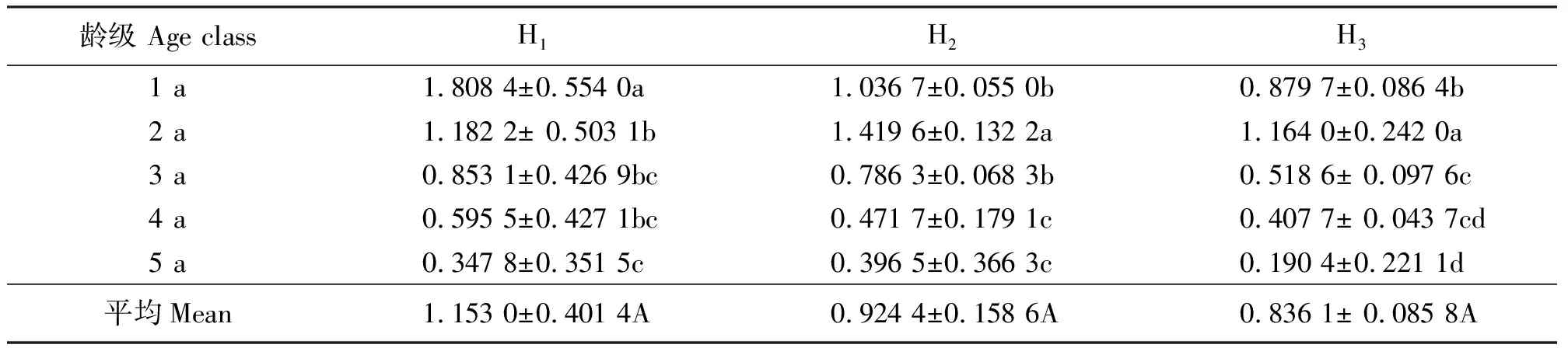
表6 不同生境假苇拂子茅种群根茎的生产力Table 6 Rhizomes’ productivity per meter of Calamagrostis pseudophragmites clonal populations in different habitats g·m-1
3 讨论与结论
本研究中,在新疆伊犁河谷地霍城县大西沟山前不同生境中,假苇拂子茅分蘖节的存活年限最高可达6 a,分蘖株最高达5个龄级,且均以5 a龄级所占比例最小;3个生境的假苇拂子茅无性系种群分蘖株在数量上均属于稳定型年龄结构。已有的研究表明,在伊犁河雅玛图段河岸、伊宁县巴依托海乡和巩乃斯河种羊场段中心岛中分布的假苇拂子茅无性系种群均为增长型种群,且最高仅达3个龄级[23]。在东北松嫩平原及内蒙古科尔沁草原区通辽县境内火电站储灰池植被恢复生境中,假苇拂子茅无性系种群分蘖株最高也为3个龄级,分蘖节最高可存活4 a,且属增长型的年龄结构[21-22]。由此可见,在新疆伊犁谷地地区,分布于沟谷适宜生境下的假苇拂子茅,由于逆温带的特殊小气候条件,其无性系种群可以达到其最大存活年限,并达到稳定结构。
禾本科多年生无性系禾草的分蘖株单株生物量代表了分蘖株的物质生产力[13]。3个生境的假苇拂子茅无性系种群均以1 a龄级分蘖株的生产力最高,且均随龄级的增加而减少,5 a龄级分蘖株的最低。在已有研究[23-24]中,分蘖株的生产力同样是以1 a龄级最旺盛。就单株生产力的整体水平而言,3个生境表现为H2>H3>H1,且生境间差异显著(P<0.05),反映了单株生产力在不同群落类型与组成间具有一定的差异。在生境H2的假苇拂子茅无性系种群中,由于没有放牧及人为干扰压力,具有旺盛的生产力,而频繁的放牧、水淹等干扰不利于其分蘖株的物质生产。因此,此结果为假苇拂子茅草地的管理、合理放牧与割草等提供了一定的理论依据。
在对伊犁河谷、松嫩平原及其他地区假苇拂子茅无性系的已有研究[21-23]中,根茎龄级均为增长型。本研究中,在霍城县山地沟谷中,假苇拂子茅无性系的根茎最多可以存活5 a。对于单位根茎的生产力,则以1 a或2 a龄级最高,这与已有研究[21-24]结果基本相同,这结果体现了不同地区、不同生境或植被类型上假苇拂子茅无性系的年轻龄级分蘖株和根茎具有旺盛的物质生产能力的共同特点。禾本科多年生草本植物的根茎是重要的繁殖器官及养分吸收与存储器官,根茎长度反映了其对地下生态空间的扩展能力,而生物量则反映根茎的养分存储能力[13]。本研究中,3个生境的假苇拂子茅根茎的生产力表现为H1>H2>H3。根据根茎龄级的划分标准,2 a根茎由1 a蘖直接供养,3 a根茎则由2 a蘖直接供养分。生境H2和H3中1、2 a分株的生物量特征恰好与根茎的相吻合,即根茎生产力的明显减弱是地上部分株旺盛的生长竞争所导致。由于生境H1中的假苇拂子茅群落郁闭度较低,尚未形成稳定的群落,加之疏松、潮湿的沙质土壤条件,无性系通过具有旺盛生命力的1 a龄级根茎向外进行扩展,因此生境H1的1 a龄级根茎具有较高的生产力。此结果为避开种子繁殖不宜进行以及植株生长缓慢等不足,实现假苇拂子茅在湿地生态工程应用中的快繁等提供了理论依据。
[1] Harper J L,White J.The demography of plants[J].Annual Review of Ecology and Systematics,1974,5(1):419-463.
[2] 刘庆,钟章成.无性系植物种群生态学研究进展及有关概念[J].生态学杂志,1995,14(3):40-45.
[3] Fischer M,van Kleunen M.On the evolution of clonal plant life histories[J].Evolutionary Ecology,2002,15:565-582.
[4] Cook R E.Clonal plant population[J].American Scientist,1983,71:244-253.
[5] Cain M L.Models of clonal growth inSoloidagoaltissima[J].Journal of Ecology,1990,78(1):27-46.
[6] Benson E J,Hartnett D C.The role of seed and vegetative reproduction in plant recruitment and demography in tall grass prairie[J].Plant Ecology,2006,187(2):163-178.
[8] Dalgleish H J,Hartnett D C.The effects of fire frequency and grazing on tall grass prairie productivity and plant composition are mediated through bud bank demography[J].Plant Ecology,2009,201(2):411-420.
[9] Klimešová J,Klimeš L.Bud banks and their role in vegetative regeneration——A literature review and proposal for simple classification and assessment[J].Perspectives in Plant Ecology, Evolution and Systematics,2007,8(3):115-129.
[10] Hendrickson J R, Briske D D.Auxiliary bud banks of two semiarid perennial grasses:Occurrence,longevity,and contribution to population persistence[J].Oecolgia,1997,110(4):584-591.
[11] 杨允菲,李建东.不同利用方式对羊草繁殖特性的影响及其草地更新的分析[J].中国草地,1994(5):34-37.
[12] 杨允菲,刘庚长,张宝田.羊草种群年龄结构及无性繁殖对策的分析[J].植物学报,1995,37(2):147-153.
[13] 李海燕,李建东,徐振国,等.内蒙古图牧吉自然保护区羊草种群营养繁殖特性的比较[J].草业学报,2011,20(5):19-25.
[14] 杨允菲,李建东,郑慧莹.松嫩平原光稃茅香无性系种群的营养繁殖特征[J].应用生态学报,1997,8(6):571-574.
[15] 张春华,杨允菲,李建东.两种碱茅种群果后营养繁殖的定量分析[J].中国草地,1996(1):31-35.
[16] 杨允菲,祝玲,李建东.松嫩平原碱化草甸星星草种群营养繁殖及有性生殖的数量特征[J].应用生态学报,1995,6(2):166-171.
[17] 朱志诚,贾东林.假苇拂子茅群落生物量初步研究[J].生态学报,1996,16(1):40-49.
[18] 邱富才,张峰,上官铁梁,等.滹沱河湿地假苇拂子茅群落生物量调查[J].山西林业科技,1998(4):26-28.
[19] 朱志诚,贾东林,岳明.黄土高原中部草地群落初级生产[J].西北大学学报(自然科学版),1998,28(6):536-539.
[20] 贺强,崔保山,赵欣胜,等.水、盐梯度下黄河三角洲湿地植物种的生态位[J].应用生态学报,2008,19(5):969-975.
[21] 杨允菲,郑慧莹.松嫩平原假苇拂子茅无性系种群的年龄结构[J].草业学报,2000,9(3):8-13.
[22] 丁雪梅,杨允菲.科尔沁草原区通辽火电厂储灰池植被恢复过程中假苇拂子茅种群构件的年龄结构[J].草业学报,2007,16(3):15-20.
[23] 赵玉,李海燕,贾娜尔,等.伊犁河谷不同生境假苇拂子茅种群构件组成及其年龄结构[J].草业学报,2009,18(2):89-94.
[24] 赵玉,李海燕,贾娜尔,等.伊犁河谷不同生境假苇拂子茅种群构件的生产力特征[J].安徽农业科学,2008,36(25):10822-10823.
[25] 杨允菲.三种拂子茅种群圆锥花序小穗分布规律的比较[J].草业学报,1997,6(3):37-41.
[26] 杨允菲,李建东.松嫩平原几种根茎型禾草种群的营养繁殖特性及其持续更新分析[J].草业学报,1996,5(2):43-48.
[27] 吴正保.杂交榛在新疆的生长适应性及抗寒性研究[D].乌鲁木齐:新疆农业大学,2008.
[28] 杨允菲,郑慧莹,李建东.根茎禾草无性系种群年龄结构的研究方法[J].东北师范大学学报(自然科学版),1998(1):49-53.
