ZNF300基因在肝癌耐药细胞HepG2/VCR中的表达及功能初探
2013-02-27李静,王涛
李 静,王 涛
(安徽医科大学基础医学院1.机能实验中心、2.病理生理学教研室,安徽合肥 230032)
目前原发性肝癌在世界范围内发病率居各种恶性肿瘤的第6位,致死率居第3位。由于肝癌早期诊断率较低,系统化疗仍是进展期肝癌病人的主要临床治疗手段[1-2]。但是肝癌病人对化疗药物的多药耐药性(multidrug resistance,MDR)成为临床治疗的一大障碍[3-4]。MDR产生的一大重要机制是肿瘤细胞内出现了耐药基因MDR-1的高表达。MDR-1基因编码的蛋白质产物为P-糖蛋白(P-glycoprotein,P-gp),它发挥的主要功能是将进入肿瘤细胞内的化疗药物泵出细胞外,降低药物的作用效率,并可以抑制 caspase的活性,减少肿瘤细胞的凋亡率[3-4]。目前许多研究组致力于P-gp抑制剂的研究工作[5-6]。然而这些研究并不能解决肝癌临床治疗的根本问题,因为许多促进肿瘤发生发展的基因,如NF-κB、ERK和HIF-1等,一方面促进肝癌细胞的增殖或抑制其凋亡,另一方面可以上调P-gp的表达促进肝癌的多药耐药性[7-9]。因此单一用药无法完全逆转肝癌的MDR,寻找新的药物作用靶点成为目前研究的热点。
ZNF300基因是一种与人胚胎发育相关的KRAB类锌指蛋白基因[10],本课题组在前期研究中发现ZNF300基因在肝癌和其他肿瘤,如结肠癌和肺癌等多种肿瘤组织中高表达,而在癌旁组织中无明显表达;进一步研究发现,ZNF300基因的过表达可以通过上调NF-κB的活性,促进ERK的磷酸化,抑制p21、p27的表达,促进肿瘤细胞的增殖和转移;通过反义cDNA的转染抑制ZNF300的表达则出现相反的效果[11-12]。由于NF-κB和ERK都可以通过上调 P-gp的高表达促进肝癌的 MDR,为观察ZNF300基因能否作为逆转肝癌多药耐药的候选药物作用靶点,在本项目中,我们拟通过体外低浓度梯度递增法构建 HepG2/VCR耐药细胞株,检测HepG2/VCR与普通HepG2细胞中的ZNF300基因与P-gp的表达差异,分析ZNF300基因与肝癌MDR的相关性;并在HepG2/VCR细胞中转染ZNF300正向或反义cDNA质粒,观察ZNF300表达上调或下调后,HepG2/VCR细胞中 P-gp的表达变化,及HepG2/VCR细胞IC50值的变化,初步分析ZNF300基因在肝癌MDR中的相关功能。
1 材料与方法
1.1 试剂和抗体人肝癌细胞株HepG2购自中科院上海细胞库;DMEM高糖培养基购自Gibco公司;胎牛血清(fatal bovine serum,FBS)购自杭州四季青公司;长春新碱(vincristine,VCR)购自合肥华泽生物有限公司;四甲基偶氮唑盐〔3-(4,5)-dimethylthiahiazo (-z-y1)-3, 5-di-phenytetrazo liumromide,MTT〕购自Sigma公司;小鼠抗人P-gp单克隆抗体购自Cell Signaling公司;小鼠抗人GAPDH抗体购自上海康成公司;兔抗人ZNF300多克隆抗体为武汉大学生命科学学院李文鑫教授馈赠;辣根过氧化物酶标记羊抗鼠第二抗体购自合肥华泽生物有限公司;ECL显影试剂盒购自Pierce公司;预染蛋白分子量Marker购自Fermentas公司;PVDF膜购自GE公司。
1.2 方法
1.2.1 细胞培养及 HepG2/VCR的构建 HepG2细胞转于含10%FBS的DMEM完全培养基中,在37℃,5%CO2饱和湿度条件下培养,每周传代2-3次,取对数生长期的细胞用于试验。肝癌多药耐药细胞株HepG2/VCR的建立采用长春新碱(VCR)低浓度梯度递增诱导法[13],直至HepG2/VCR细胞能在含VCR 1 mg·L-1的培养液中维持生长,并利用MTT法检测其耐药性。
1.2.2 HepG2/VCR耐药性的鉴定 为鉴定HepG2/VCR的耐药性,将 HepG2/VCR和普通HepG2细胞培养至指数生长期后,按5×103个细胞/孔接种于96孔板,培养24 h后,加入不同浓度VCR(0.01、0.1、1、10 和100 mg·L-1)处理24 h,进行MTT检测,绘制生长曲线。
1.2.3 质粒构建和转染 正向和反向ZNF300基因cDNA克隆入IRES-GFP真核表达质粒中,分别命名为 IRES-ZNF300-GFP和 IRES-ASZNF300-GFP,克隆方法见参考文献[12]。用Lipofectamine 2000转染试剂将IRES-ZNF300-GFP和IRES-ASZNF300-GFP及其对照质粒IRES-GFP转染入HepG2/VCR细胞中,转染方法为:将HepG2/VCR细胞按5×105个细胞/孔接种于6孔板,在含5%CO2、37℃恒温培养箱中培养,直至转染前细胞密度达到80% ~90%;按转染试剂说明书配制转染复合物(Lipofectamine 2000):在250 μl Opti-MEM 中加入10 μl转染试剂;质粒复合物:在250 μl Opti-MEM中加入4 μg质粒;室温静置5 min后将质粒复合物加入转染试剂复合物中,使之形成DNA-脂质体复合物,室温静置20 min,每孔细胞中加入1.5 ml无血清无双抗DMEM培养液,将DNA-脂质体复合物轻轻混匀加入细胞中,在含5%CO2、37℃恒温培养箱中培养5 h后;将每孔细胞更换成2 ml有血清的DMEM培养液。转染24 h后进行后续实验。
1.2.4 Western blot检测 Western blot方法用于检测HepG2/VCR和普通HepG2细胞中ZNF300和P-gp蛋白质的表达差异,及HepG2/VCR质粒转染后ZNF300和P-gp蛋白质的表达变化。收取经上述细胞,用含有蛋白酶抑制剂、磷酸酶抑制剂、氟化钠和钒酸钠的加样缓冲液冰上静置裂解细胞10 min,再100℃变性10 min即为全细胞裂解液,SDS-PAGE凝胶电泳后冰浴湿式转印至PVDF膜,参数为恒压80V ×3 h.用含 0.5 g·L-1脱脂奶粉和 0.5 ml吐温20的PBST封闭30 min后,按抗体厂家建议稀释一抗后4℃敷膜结合过夜,待检测抗体为ZNF300多克隆抗体和P-gp单克隆抗体,以GAPDH抗体为内参。一抗结合后PBS-T洗膜10 min×3次,加入适当稀释度的二抗,室温揺床结合2 h,再次PBS-T洗膜10 min×3次,暗室内ECL显色,柯达胶片感光显影并扫描。
1.2.5 MTT检测及IC50值计算 HepG2/VCR细胞按5×103个细胞/孔接种于6孔板,进行ZNF300相关质粒转染后48 h,用0.25%胰酶消化重悬,按5×103个细胞/孔接种于96孔板,24 h后加入不同浓度VCR(0.125、0.25、0.5、1和2mg·L-1)干预 24 h,在终止培养前4 h加MTT溶液(0.5 mg·L-1)。培养结束后弃上清,每孔加入100 μl DMSO,置于摇床上15 min,使结晶物充分溶解;置酶标仪上检测各孔吸光度值(A570nm),实验至少重复3次。根据MTT结果绘制生长曲线,并采用中效分析软件(Logit法)计算质粒转染后,HepG2/VCR细胞对VCR的半数抑制浓度值(50%inhibitory concent ration,IC50)变化,按以下公式计算耐药指数(resistance index,RI)。RI=IC50质粒转染后的细胞/IC50亲本细胞。
1.2.6 统计学处理 采用SPSS 13.0统计软件对实验数据进行分析,计量资料以±s表示,组间数据的显著性检验采用ANOVA法分析。
2 结果
2.1 肝癌耐药细胞株HepG2/VCR耐药性的鉴定筛选出的HepG2/VCR细胞和HepG2细胞在其指数生长期时,分别加入不同浓度 VCR(0.01、0.1、1、10和100 mg·L-1)处理24 h,经MTT检测后绘制生长曲线后发现,HepG2/VCR细胞对VCR的耐药性相对于 HepG2细胞明显增高(P<0.05,ANVOA)(Fig 1),说明肝癌耐药细胞株HepG2/VCR构建成功。

Fig 1 MTT analysis of HepG2/VCR and HepG2 treated with different concentrations of VCR
2.2 Western blot检测发现HepG2/VCR细胞中ZNF300和P-gp表达增高HepG2/VCR细胞和HepG2细胞在其指数生长期时收取蛋白质,进行Western blot检测,样品各点两道,用兔抗人ZNF300多克隆抗体和鼠抗人P-gp单克隆抗体进行检测,以GAPDH作为内参,结果如Fig 2所示,可见在GAPDH表达基本一致的情况下,HepG2/VCR细胞中ZNF300和P-gp的表达明显高于HepG2细胞,提示ZNF300基因与HepG2/VCR细胞的耐药性和P-gp的表达有相关性。
2.3 ZNF300基因表达变化对HepG2/VCR细胞中P-gp表达及耐药性的影响在HepG2/VCR细胞中转染,将IRES-ZNF300-GFP和IRES-ASZNF300-GFP及其对照质粒IRES-GFP,转染后48 h,收取蛋白质,用兔抗人ZNF300多克隆抗体和鼠抗人P-gp单克隆抗体进行Western blot检测,以GAPDH作为内参,结果如Fig 3所示,可见在GAPDH表达基本一致的情况下,IRES-ZNF300-GFP组细胞中ZNF300和P-gp表达明显高于空载体组和亲本细胞组,IRES-ASZNF300-GFP组细胞中结果相反,说明质粒转染后可有效促进ZNF300在细胞中的高表达或Knockdown,ZNF300高表达可上调 P-gp的表达,ZNF300基因knockdown则可抑制P-gp的表达。质粒转染后,加入不同浓度 VCR(0.125、0.25、0.5、1和2 mg·L-1)干预24 h行MTT检测绘制生长曲线(Fig 4),并计算 IC50值和 RI值变化(Tab 1),可见ZNF300基因高表达可促进HepG2/VCR的耐药性,而ZNF300基因Knockdown则有相反的结果(P<0.05)。说明ZNF300基因可以通过上调P-gp的表达促进HepG2/VCR的耐药性。

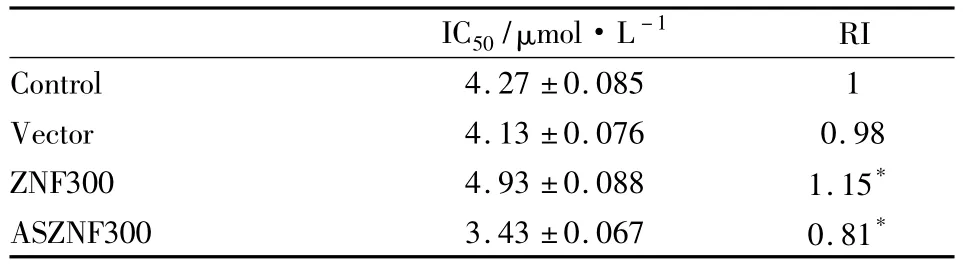
Tab 1 Drug resistance of HepG2/VCR improved by ZNF300 overexpression
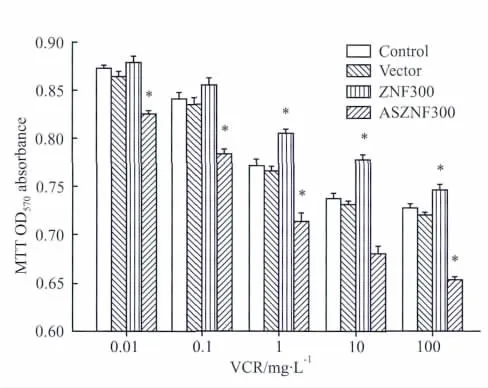
Fig 4 Drug resistance of HepG2/VCR cells promoted by ZNF300 overexpression
3 讨论
对于原发性肝癌,有效的临床治疗手段在于早期诊断和手术切除原发病灶,但是目前临床上确诊的大多数肝癌病人已处于进展期,所以系统化疗仍是目前肝癌临床治疗的主要手段[1-2]。然而肝癌病人对传统化疗药物均会出现多药耐药(MDR)现象,所以肝癌病人的预后往往极为不好[3-4]。近年来随着分子靶向药物的研究进展,以Sorafenib为代表的一些药物被批准上市,给肝癌的临床治疗带来新的希望,然而现在发现病人同样会对Sorafenib产生耐药性[3]。所以对于肝癌MDR逆转药物或药物作用靶点的研究成为目前的研究热点。
现在认为肝癌MDR产生的一个最主要的机制在于 MDR-1基因产物 P-糖蛋白(P-gp)的高表达[4]。P-gp是一个170 ku大小的蛋白质成分,表达于肿瘤细胞的细胞膜,属于ATP结合盒转运蛋白(ATP-binding cassette transporter)家族成员,主要功能在于将进入细胞的药物泵出细胞外以达到减少药物对细胞毒性的目的[4]。在本项目中,我们采用长春新碱(VCR)低浓度梯度递增诱导法建立HepG2/VCR耐药细胞株,通过MTT检测确定其构建成功,经Western blot检测后发现,HepG2/VCR细胞中的P-gp表达水平明显高于HepG2细胞,这与前期研究报道相符[13]。
然而目前许多抑制P-gp表达的药物并不能根本解决肝癌MDR的实际问题,因为肝癌MDR的产生是一个多因素参与的过程,现在发现许多促进肿瘤发生发展的基因,如NF-κB和ERK,一方面可以抑制肿瘤细胞凋亡,另一方面亦可上调P-gp的表达,在肝癌 MDR的产生过程中发挥重要的作用[4,7-8]。所以寻找新的药物作用靶点与P-gp抑制剂共同作用来逆转MDR成为一种新的策略。
ZNF300基因最初是从人胚胎cDNA文库中筛选出来的一种KRAB类锌指蛋白基因,前期研究发现其KRAB区可以发挥转录抑制作用,其锌指区可以结合CG富含的启动子区域[10-12]。本课题组前期研究发现ZNF300基因可以通过调控TRAF2基因的表达,同时结合IKKβ蛋白质来促进NF-κB通路的活性,进一步研究发现ZNF300高表达可以促进ERK的磷酸化,从而促进肿瘤细胞的增殖和转移,而NF-κB和ERK都可上调 P-gp的表达[12]。利用免疫组织化学检测我们发现ZNF300基因在肝癌等多种肿瘤组织细胞中高表达,而在癌旁组织中表达较低[12]。综合上述研究基础,在本项目中,我们利用Western blot检测发现HepG2/VCR细胞中ZNF300的表达明显高于HepG2细胞,并与P-gp的表达有相关性,提示ZNF300基因可能参与肝癌MDR的形成过程。在进一步的实验中,我们将正向和反向ZNF300基因cDNA转染入HepG2/VCR细胞中,经Western blot检测发现正向cDNA的转染可以明显促进ZNF300的表达,并上调P-gp在HepG2/VCR中的表达,MTT检测发现ZNF300基因高表达可以促进HepG2/VCR的耐药性。而反向ZNF300基因cDNA则有完全相反的结果。
上述实验结果说明ZNF300基因在肝癌耐药细胞株中高表达,并可以通过上调耐药基因P-gp的表达促进肝癌的多药耐药性,初步证明ZNF300可以作为逆转肝癌MDR的潜在药物作用靶点。而ZNF300基因在肝癌MDR发生发展过程中的发挥作用的具体机制尚待进一步研究。
[1] El-Serag H B.Hepatocellular carcinoma[J].N Engl J Med,2011,365(12):1118-27.
[2] Livraghi T,Mäkisalo H,Line P D.Treatment options in hepatocellular carcinoma today[J].Scand J Surg,2011,100(1):22-9.
[3] Gauthier A,Ho M.Role of sorafenib in the treatment of advanced hepatocellular carcinoma:An update[J].Hepatol Res,2013,43(2):147-54.
[4] D'Alessandro N,Poma P,Montalto G.Multifactorial nature of hepatocellular carcinoma drug resistance:could plant polyphenols be helpful[J]?World J Gastroenterol,2007,13(14):2037-43.
[5] 戴春岭,符立梧.肿瘤多药耐药逆转剂的研究进展[J].中国药理学通报,2005,21(5):513-8.
[5] Dai C L,Fu L W.The development of the reversal of the tumor multidrug resistance[J].Chin Pharmacol Bull,2005,21(5):513-8.
[6] 石俊敏,张冬梅,姚 楠,等.三萜皂苷Saxifragifolin D抑制人肝癌耐药细胞HepG2/ADM生长并诱导细胞凋亡作用研究[J].中国药理学通报,2012,28(1):34-8.
[6] Shi J M,Zhang D M,Yao N,et al.Studies of the cell proliferative inhibition and apoptosis induced by Saxifragifolin D on HepG2/ADM cells[J].Chin Pharmacol Bull,2012,28(1):34-8.
[7] Liu Y,Lou G,Wu W,et al.Involvement of the NF-κB pathway in multidrug resistance induced by HBx in a hepatoma cell line[J].J Viral Hepat,2011,18(10):e439-46.
[8] Yan F,Wang X M,Pan C,et al.Down-regulation of extracellular signal-regulated kinase 1/2 activity in P-glycoprotein-mediated multidrug resistant hepatocellular carcinoma cells[J].World J Gastroenterol,2009,15(12):1443-51.
[9] Comerford K M,Wallace T J,Karhausen J,et al.Hypoxia-inducible factor-1-dependent regulation of the multidrug resistance(MDR1)gene[J].Cancer Res,2002,62(12):3387-94.
[10] Gou D,Wang J,Gao L,et al.Identification and functional analysis of a novel human KRAB/C2H2 zinc finger gene ZNF300 [J].Biochim Biophys Acta,2004,1676(2):203-9.
[11] Xu J H,Wang T,Wang X G,et al.PU.1 can regulate the ZNF300 promoter in APL-derived promyelocytes HL-60[J].Leuk Res,2010,34(12):1636-46.
[12] Wang T,Wang X G,Xu J H,et al.Overexpression of the human ZNF300 gene enhances growth and metastasis of cancer cells through activating NF-κB pathway[J].J Cell Mol Med,2012,16(5):1134-45.
[13]彭仕芳,傅 蕾,谭德明.热休克蛋白27高表达与肝细胞癌耐药相关[J].中华肝脏病杂志,2007,15(5):362-5.
[13] Peng S F,Fu L,Tan D M.Heat-shock protein 27 linked to multidrug resistance in human hepatic cancer cell lines[J].J Chin Liver Dis,2007,15(5):362-5.