乙酸对奶牛乳腺上皮细胞活力及CD36和FABP3基因表达的影响
2013-02-20齐利枝闫素梅赵艳丽
■齐利枝 闫素梅 生 冉 赵艳丽 金 鹿
(内蒙古农业大学动物科学学院,内蒙古呼和浩特 010018)
乳腺中约50%的脂肪酸来源于乙酸等乳脂前体物在乳腺内的从头合成,主要包括中短链脂肪酸(C4∶0~C14∶0)及50%的C16∶0;长链脂肪酸(C18∶0及50%的C16∶0)除油酸是乳腺中由硬脂酸通过△9-去饱和酶系统产生外,其余均来自日粮[1]。乳脂是最容易受到遗传、生理和日粮等因素影响而发生改变的乳成分。研究已经证实乙酸等用于从头合成的乳脂前体物与外源供给的长链脂肪酸(LCFA)对乳脂肪的合成有直接的影响,日粮中增加LCFA的供给对脂肪酸的从头合成有显著的抑制作用,增加中、短链脂肪酸(SMCFA)的供给可以减少用于从头合成的短链脂肪酸(SCFA),可提高乳脂含量,改变其脂肪酸组成[2]。因此,深入探讨乳脂前体物对乳脂肪合成的影响机理对不断改善乳脂组成及乳品质有重要意义。乙酸作为奶牛乳腺脂肪酸内源合成的主要前体物,体外研究结果表明,其可以促进奶牛乳腺上皮细胞甘油三酯(TAG)合成。孔庆洋等[3]研究了不同浓度的乙酸钠对奶牛乳腺上皮细胞胞外TAG浓度的影响,结果表明,随着乙酸添加剂量的增加,TAG浓度显著增加。Maxin等(2011)[4]的研究表明,瘤胃灌注乙酸处理组乳脂率增加了6.5%,而且改变了乳脂组成。提示乙酸对乳脂合成有一定的影响,但其机制仍然不清楚。分化抗原簇36(CD36)参与乳脂分泌,但其更重要的作用是转运脂肪酸进入奶牛乳腺细胞(Bionaz等,2008a)[5]。脂肪酸结合蛋白3(FABP3)参与细胞内脂肪酸的转运,并且在泌乳期奶牛乳腺中的FABP3 mRNA的相对表达量上调(Bionaz和Loor,2008b)[6]。因此,乙酸对乳脂肪合成的影响可能与CD36和FABP3 mRNA的基因表达有关,而关于这方面的研究较少。本试验主要研究乙酸对奶牛乳腺上皮细胞活力及CD36和FABP3 mRNA相对表达量的影响,为进一步研究乙酸对乳脂肪合成的影响提供理论依据。
1 材料与方法
1.1 试剂与仪器
DMEM/FA12培养基(Gibco,美国)、胎牛血清(FBS,Gibco,美国)、无脂肪酸牛血清白蛋白(BSA,Equitech-Bio,美国)、催乳素(Gibco,美国)、表皮生长因子(Gibco,美国)、胰蛋白酶(Gibco,美国)、胰岛素转铁蛋白(Gibco,美国)、双抗(Gibco,美国)、乙二胺四乙酸(EDTA,Gibco,美国)、氢化可的松(Sig⁃ma,美国)、噻唑蓝(MTT,Sigma,美国)、二甲基亚砜(DMSO,Sigma,美国)、乙酸钠(Sigma,美国)、RNAprep pure Cell/Bacteria Kit(TIANGEN,北京)、PrimeScript RT Master Mix(TaKaRa,大连)和 SYBR Premix Ex TaqTMⅡ(TaKaRa,大连)。实时定量PCR仪(Roche,瑞士)、酶标仪(Biotek,美国)、倒置显微镜(Olympuse,日本)。
1.2 乳腺上皮细胞的培养
采用胶原酶消化法获得乳腺上皮细胞。选取健康的荷斯坦奶牛,取其乳腺组织,去除组织外层,剪取若干约1 cm3的组织块,置于预冷的PBS溶液中。在无菌超净台中将组织块用PBS洗净后,去除组织块表层将其剪成糊状。加入0.5%胶原酶Ⅱ溶液,置于37℃和5%CO2的培养箱中消化1 h,每20 min轻摇离心管。然后用孔径80目的细胞滤网进行过滤,收集细胞滤液,1 500 r/min离心5 min,弃掉上清液,加入含有10%FBS的DMEM/F12细胞培养液,用移液枪吹打均匀,接种在25 cm2培养瓶中,置于37℃和5%CO2的培养箱中培养。当细胞的融合度为80%~90%时,用胰蛋白酶消化处理乳腺上皮细胞并进行传代。本试验采用第3代细胞。
1.3 试验设计
将传第3代的乳腺上皮细胞悬液接种于细胞培养板上,每孔加入含10%FBS的DMEM/F12培养液,于 37℃、5%CO2培养箱培养48 h。将培养48 h的奶牛乳腺上皮细胞培养孔随机分为1个对照组和5个试验组。每组分别加入含0、4、6、8、10、12 mmol/l乙酸(以乙酸钠的形式添加)的DMEM/F12培养液,并用1 g/l无脂肪酸的BSA替代培养液中的FBS,其中0 mmol/l为对照组。置于37℃和5%CO2的培养箱中继续培养48 h。
1.4 测试指标与方法
1.4.1 细胞活力
细胞活力的检测采用MTT法,以细胞相对增殖率(Relative Growth Rate,RGR)表示。在细胞培养结束前4 h,每孔加入20 μl MTT(5 mg/ml);4 h后弃上清液,每孔加入100 μl DMSO,振荡10 min后,用全自动酶标仪(Synergy H4 Reader)在490 nm波长下检测各培养孔的吸光值(OD490)。RGR=(试验组OD490/对照组OD490)×100%。每个处理4个重复。
1.4.2 乳腺上皮细胞内FABP3和CD36 mRNA的表达
乳腺上皮细胞内FABP3和CD36 mRNA相对表达量采用实时荧光定量PCR法检测。将每孔1×105细胞悬浮液接种于6孔培养板,培养48 h,在每孔加入含有不同浓度乙酸的诱导培养基中继续培养48 h后,提取RNA。细胞RNA的提取、反转录和荧光定量PCR分别采用RNAprep pure Cell/Bacteria Kit试剂盒、PrimeScript®RT reagent Kit和SYBR Premix Ex TaqTMⅡ试剂盒进行操作。选用管家基因磷酸甘油醛脱氢酶(GAPDH)作为内参基因,引物见表1。实时荧光定量PCR的反应程序为:95.0 ℃、30 s;95.0 ℃、30 s,Tm 30 s,72.0 ℃20 s,40cycles;72 ℃、7 min;溶解曲线程序为:70~95℃,每6 s升高0.5℃,51cycles。每个处理组6个重复。采用2-△△Ct法进行相对定量数据分析[7]。
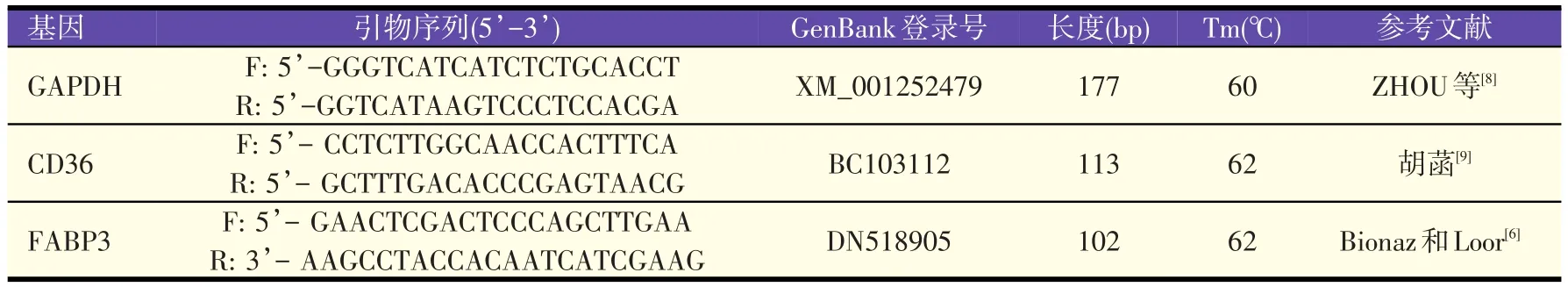
表1 Primer sequence
1.5 数据处理
试验数据采用SAS(SAS 9.0)软件的回归统计程序进行,对不同浓度的乙酸处理效应进行一次线性和二次曲线回归分析,P<0.05表示差异显著,
0.05<P<0.10表示差异趋于显著。
2 结果与分析
2.1 添加乙酸对奶牛乳腺上皮细胞活力的影响(见表2)
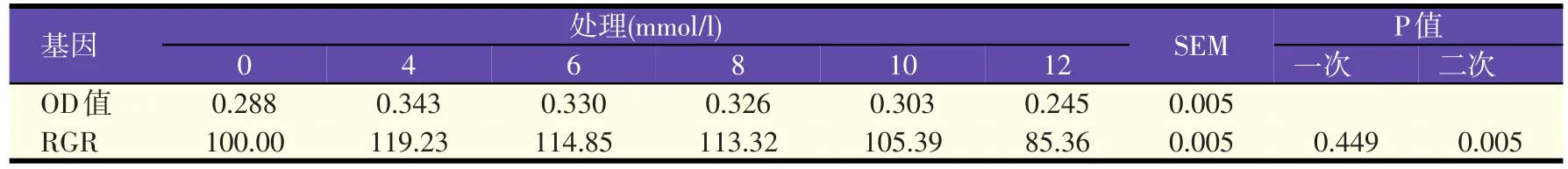
表2 乙酸对奶牛乳腺上皮细胞活力的影响
如表2所示,4~10 mmol/l的所有乙酸处理组RGR均高于对照组,其中,以4~8 mmol/l乙酸处理组较高。当乙酸浓度增加到12 mmol/l时,RGR降低,并低于对照组。回归分析结果表明,随着乙酸浓度的增加RGR呈二次曲线增加,二者呈显著的剂量依赖关系(P=0.005),回归方程为y=-0.006 85x2+0.071 15x+1.000 48,R2=0.970 6;其中x为乙酸添加水平,y为RGR。这些结果提示低浓度的乙酸可促进奶牛乳腺上皮细胞活力,而高浓度则表现出抑制作用。
2.2 添加乙酸对奶牛乳腺上皮细胞FABP3和CD36 mRNA相对表达量的影响(见表3)

表3 乙酸对奶牛乳腺上皮细胞CD36和FABP3 mRNA相对表达量的影响
从表3可以看出,添加不同浓度的乙酸均下调了CD36 mRNA相对表达量,且以8~12 mmol/l乙酸组CD36的表达量较低。FABP3表达量的结果正好相反,即4~12 mmol/l的所有乙酸处理组FABP3基因表达量均不同程度高于对照组,其中以10~12 mmol/l乙酸组表达量较高。从回归统计结果看,CD36的相对表达量随着乙酸添加浓度的增加呈显著的一次线性(P=0.011)和二次曲线(P=0.000)降低,回归方程为:y=-0.038 59x+0.903 89,R2=0.831 0;y=0.004 55x2-0.093 14x+1.122 93,R2=0.994 82;其中x为乙酸添加水平,y为CD36 mRNA相对表达量。随着乙酸添加浓度的增加,FABP3的表达量呈显著的一次线性(P=0.001)和二次曲线(P=0.010)增加,回归方程为:y=0.247 44x+0.98 84,R2=0.945 0;y=0.006 75x2+0.166 38x+1.122 93,R2=0.955 2;其中X为乙酸添加水平,y为FABP3 mRNA相对表达量。说明乙酸的添加可下调CD36的基因表达量,上调FABP3表达量。
3 讨论
细胞活力是用来反映细胞存活和增殖的指标。本研究结果发现,乙酸作为脂肪酸内源合成的主要前体物和主要能源物质,对奶牛乳腺上皮细胞活力有一定的调节作用,二者呈剂量依赖关系,低浓度的乙酸可促进奶牛乳腺上皮细胞活力,而高浓度则表现出抑制作用。目前关于乙酸对奶牛乳腺上皮细胞活力的研究尚未见报道,其机理还需要进一步探讨。
FABP和CD36是奶牛乳腺中两种主要的LCFA转运蛋白,在牛乳腺中FABP和CD36的共表达提示这两种蛋白存在相近的功能关系(Spitsberg等,1995)[10]。在奶牛乳腺组织中FABP3、FABP4和FABP5三种异构体的mRNA表达量较高,在泌乳阶段FABP3 mRNA表达量最高(Bionaz和Loor,2008b)[6]。FABP3参与细胞内LCFA的转运,CD36主要将脂肪酸转运进入乳腺细胞,二者与乳腺上皮细胞对长链脂肪酸的摄取与转运有关。然而,目前关于乙酸对FABP3和CD36基因表达的研究极少。Yonezawa等(2004)在奶牛乳腺上皮细胞培养液中分别添加10 mmol/l乙酸、丁酸或辛酸,研究结果表明,辛酸显著上调了细胞中CD36 mRNA表达量,但乙酸和丁酸对CD36表达量的促进作用远低于辛酸,并提示乙酸、丁酸或辛酸CD36表达量的上调作用似乎与碳原子的数量有关[11]。Storry等(1965)[12]体内试验的研究结果表明,瘤胃灌注乙酸增加了乳脂中C4-C16脂肪酸的量,而降低了所有C18脂肪酸的量。崔海(2011)[2]的研究结果也表明,在奶牛日粮中添加SMCFA,乳脂中C10∶0、C12∶0、C14∶0、C14∶1、C16∶0和C14∶1含量增加,除了c9-C18∶1外其它C18脂肪酸含量均增加。这些结果提示,外源补充乙酸钠降低了乳脂中长链脂肪酸的量,即降低了乳脂中来源于外源供给的LCFA的比例。本试验的研究结果表明,不同浓度的乙酸处理组均下调了CD36 mRNA相对表达量。这一结果可能解释了外源补充乙酸钠是通过下调脂肪酸转运蛋白CD36 mRNA表达量,影响乳腺上皮细胞对LCFA的摄取进而引起乳脂中LCFA的合成受限。然而,本试验的结果也表明,FABP3的mRNA表达量随着乙酸添加浓度的增加而增加,与CD36的表达量结果相反。FABP3和CD36尽管都是LCFA的转运蛋白,但前者主要与细胞内的转运有关,后者与细胞的摄取有关,其功能并不相同,因此乙酸钠对这两种转运蛋白基因表达量的调节作用并不矛盾。目前关于该领域的研究甚少,还需要进一步探讨。
4 结论
①乳腺上皮细胞活力与乙酸浓度呈显著的二次剂量依赖关系,低浓度的乙酸可促进奶牛乳腺上皮细胞活力,而高浓度则表现出抑制作用。
②不同浓度的乙酸处理均下调了CD36 mRNA相对表达量,上调了FABP3 mRNA表达量。
