宇佐美曲霉阿魏酸酯酶A基因的克隆表达及酶学性质研究
2013-02-19龚燕燕邬敏辰
龚燕燕, 殷 欣, 邬敏辰, 曾 妍
(1.江南大学 药学院,江苏 无锡 214122;2.江南大学 工业生物技术教育部重点实验室,江苏 无锡 214122;3.江南大学 无锡医学院,江苏 无锡214122)
阿魏酸(Ferulic acid),化学名为 4-羟基-3-甲氧基苯丙烯酸,是普遍存在于多种植物中的一种酚酸[1],除少数以游离形式存在外,在细胞壁中阿魏酸或二聚体阿魏酸主要以酯键的形式连接在阿拉伯木聚糖的阿拉伯糖残基上[2-3],可增强阿拉伯木聚糖链的强度,但从空间上限制了动物和微生物对植物细胞壁中纤维素、半纤维素的有效降解。
阿魏酸酯酶(E.C.3.1.1.73,Ferulic acid esterase,FAE)又称肉桂酸酯酶,是羧酸水解酶的一个亚类,属于胞外酶,能水解阿魏酸甲酯、低聚糖阿魏酸酯和多糖阿魏酸酯中的酯键,将阿魏酸释放出来[4-5]。自1991年Faulds[6]等首次分离出阿魏酸酯酶以来,已有超过30种阿魏酸酯酶被纯化,其主要生物功能是水解植物细胞壁中多糖与阿魏酸连结的酯键,释放出游离的单体阿魏酸或阿魏酸二聚体[7]。阿魏酸因其具有较强的抗氧化活性和防腐作用而被广泛应用,它也是目前诸多药材中的有效成分,并在新型药材开发和化妆品行业广受青睐[8]。反式阿魏酸在美国、日本已被允许用作食品添加剂[1],合成品阿魏酸钠具有治疗偏头痛、抑制花生四烯酸代谢等作用[9]。目前生产阿魏酸的方法有化学合成法、碱法和酶法,化学合成法因为反应周期长和产率低限制了其发展;碱解法是目前商品化生产阿魏酸的主要途径,但该法受原料限制,产量低,成本高,因而国际市场上阿魏酸的价格居高不下[10]。近年来通过酶解法生产阿魏酸已逐渐吸引众多学者的关注。已有报道表明,阿魏酸酯酶可以和其它的半纤维素酶(如木聚糖酶)协同作用促使微生物对植物细胞壁进行最大程度的降解[11]。因此,阿魏酸酯酶在造纸业、饲料业、乙醇制造等诸多领域具有广泛的应用前景。
作者利用实验室保藏的宇佐美曲霉菌株Aspergillus usamii E001,结合生物信息学分析的手段,借助RT-PCR技术克隆了编码阿魏酸酯酶A的cDNA片段,并实现了其在毕赤酵母GS115中的分泌表达。同时,对reAusFaeA的酶学性质进行了初步研究,优良的酶学性质表明该酶具有较大的应用潜力。
1 材料与方法
1.1 菌株和质粒
宇佐美曲霉(Aspergillus usamii)E001菌株由江南大学无锡医学院分子生物学实验室筛选和保藏;E.coli JM109、E.coli DH5α 为克隆宿主菌,pUCm-T为克隆质粒,购于上海Sangon公司;P.pastoris GS115为表达宿主菌,pPIC9K表达质粒,购自Invitrogen公司。
1.2 工具酶和试剂
各种限制性内切酶、rTaq DNA聚合酶、T4 DNA连接酶、250 bp DNA Ladder Marker和低分子量蛋白质Marker均购自TaKaRa公司;YNB、生物素、酵母粉、 蛋白胨、G418、UNIQ-10柱式 Trizol总 RNA抽提试剂盒和EZ-10 Spin Column DNA Gel Extraction Kit购自上海Sangon公司;标准反式阿魏酸与阿魏酸甲酯购自盐城朗德化学有限公司;其他试剂均为国产或进口分析纯。
1.3 培养基
种子活化培养基(质量分数):2%玉米粉,3%豆饼粉,3%KH2PO4,1%CaCl2,1%MgSO4;诱导培养基(用于提 RNA):0.9%NaNO3,0.05%KH2PO4,0.05%MgSO4,0.4%酵 母 提 取 物 ,20%麸 皮 提 取 液 ;LB、YPD、MD、BMGY和BMMY培养基参考Multi-Copy Pichia Expression Kit(Invitrogen公司)操作手册。
1.4 引物设计及合成
基于宇佐美曲霉与黑曲霉基因序列的相似性[12-13],参照NCBI上公布的Aspergillus niger CBS 513.88菌株基因组中阿魏酸酯酶A的基因序列,结合生物信息学分析的手段,设计出一对扩增编码AusFaeA成熟肽cDNA的特异性上下游引物。

1.5 宇佐美曲霉总RNA的提取
将A.usamii E001接种至种子活化培养基中,于30℃、220 r/min条件下振荡培养24 h,以3%的接种量转至诱导培养基中,于上述条件下振荡培养24 h后收集菌体,用UNIQ-10柱式Trizol总RNA抽提试剂盒提取A.usamii的总RNA。
1.6 编码AusFaeA成熟肽cDNA的克隆及分析
以提取的A.usamii总RNA为模板、Oligo dTAdaptor为引物,逆转录合成cDNA第一链。以其为模板、Fae-F和M13 primer M4为引物进行第一轮PCR 扩增,反应体系为 50 μL,包括:5×PCR 缓冲液10 μL, 逆转录产物 2 μL,dNTP (2.5 mmol/L) 1.5 μL,Fae-F、M13 Primer M4 各 0.5 μL,rTaq 酶 0.25 μL,双蒸水 35.25 μL;反应条件为:94 ℃ 5 min,30个循环(94 ℃ 30 s,51 ℃ 30 s,72 ℃ 60 s),72 ℃ 10 min。以第一步PCR产物为模板、Fae-F和Fae-R为引物进行第二轮PCR扩增,反应条件为:94℃5 min,30 个循环(94 ℃ 30 s,53 ℃ 30 s,72 ℃ 60 s),72℃10 min。PCR产物经1%琼脂糖凝胶电泳后,割胶回收目的条带,具体操作参见EZ-10 Spin Column DNA Gel Extraction Kit说明书。将纯化的PCR产物直接与pUCm-T连接,转化JM109感受态细胞;经蓝白斑初选、质量分数0.7%琼脂糖凝胶电泳和PCR鉴定正确后,送上海Sangon公司测序,测序正确的重组质粒命名为pUCm-T-AusfaeA。最后,将测得的序列在NCBI上进行Blast分析。
1.7 重组表达质粒的构建
将测序验证过的pUCm-T-AusfaeA用EcoRⅠ和NotⅠ双酶切,割胶回收目的基因AusfaeA,并与经同样双酶切的pPIC9K质粒连接,转化DH5α感受态细胞,经PCR筛选鉴定获重组表达质粒pPIC9K-AusfaeA后,送上海Sangon公司测序。
1.8 AusfaeA在毕赤酵母GS115中的表达
pPIC9K和测序验证过的pPIC9K-AusfaeA经SalⅠ线性化后,用电穿孔法分别将其导入毕赤酵母GS115中,涂布于MD平板上筛选重组毕赤酵母;将在MD平板上生长良好的菌落用牙签点种至含不同浓度G418的YPD平板上筛选抗最高浓度2.0 mg/mL G418的重组毕赤酵母,分别命名为GS115/9K和GS115/AusfaeA;参照《分子克隆实验指南》的方法提取重组GS115基因组DNA,利用通用引物5'-AOX和3'-AOX PCR鉴定目的基因是否整合入GS115基因组内。GS115/9K和GS115/AusfaeA的常规表达参见Multi-Copy Pichia Expression Kit(Invitrogen公司)操作手册。
1.9 reAusFaeA的分离纯化与活性测定
将诱导表达的发酵液经8 000 r/min离心5 min后收集上清液,再经质量分数80%饱和度(NH4)2SO4盐析,离心收集沉淀,复溶于适量20 mmol/L pH 6.0磷酸氢二钠-磷酸二氢钠缓冲液,经透析、超滤浓缩(超滤膜的截留相对分子质量为10 000,Millipore公司)后用Sephadex G-75凝胶过滤层析柱纯化。蛋白质浓度测定采用Bradford法[14],以牛血清白蛋白为标准。采用SDS-PAGE对表达产物进行分析。
以阿魏酸甲酯(MFA)为底物,用高效液相色谱法(HPLC)测定经reAusFaeA水解后的产物中阿魏酸的释放量。具体步骤按zhang[15]提供的方法稍作修改:移取 900 μL MFA(1 mM,pH 6.0 磷酸盐缓冲液配制)于2 mL EP管中,45℃保温10 min,加入100 μL适当稀释的酶液,反应10 min后加入400 μL冰乙酸终止反应,立即混匀进行高效液相色谱分析。以预先在酶溶液中加入400 μL冰乙酸,再加底物溶液的反应物为对照。在测定条件下 (45℃、pH 6.0),以每分钟产生1 μmol阿魏酸所需的酶量为一个酶活单位(U)。
1.10 reAusFaeA酶学性质的测定
1.10.1 reAusFaeA的最适温度及温度稳定性 在pH 6.0的条件下反应10 min,测定reAusFaeA在不同温度下(30~70℃)的酶活性。以某一温度下的最高酶活性为100%,其他温度下酶活性与最高酶活性的比值为该温度下的相对酶活,绘制温度-相对酶活性曲线。将酶液分别置于45、50、55℃下处理10、20、30、40、50、60 min,收取酶液,采用高效液相色谱法测定残余酶活性。以0 min分钟取出酶液的酶活性为100%,残余酶活与其比值为相对酶活,绘制时间-相对酶活性曲线。
1.10.2 reAusFaeA的最适pH及pH稳定性 在最适反应温度下反应10 min,测定reAusFaeA在不同pH值(pH 3.5~7.5的磷酸氢二钠-柠檬酸缓冲液)下的酶活性。以某一pH值下的最高酶活性为100%,其他pH下酶活性与最高酶活性的比值为该pH下的相对酶活,绘制pH-相对酶活性曲线。将用不同pH值缓冲液 (pH 3.5~7.5的磷酸氢二钠-柠檬酸缓冲液)稀释25倍的酶液于35℃保温1 h,按标准测定方法测定残余酶活性。以其中酶活性最高者为100%,其他pH下酶活性与最高酶活性的比值为该pH下的相对酶活,绘制pH-相对酶活性曲线。
1.10.3 金属离子和EDTA对reAusFaeA酶活性的影响 将酶液与不同金属离子或EDTA(终浓度为2.0 mmol/L)混匀后于35℃保温1 h,按标准测定方法测定残余酶活性。以只加缓冲液同样经过35℃保温1 h的酶活性为100%,来衡量金属离子和EDTA对reAusFaeA酶活性的影响。
2 结果与分析
2.1 编码AusFaeA成熟肽cDNA的克隆及分析
按照1.6中的方法进行RT-PCR及PCR反应,经质量分数1%琼脂糖凝胶电泳检测得到的电泳图如图1所示,目的条带位于800 bp左右的位置,与预期的基因片段大小相符。将目的条带割胶回收后与pUCm-T质粒进行连接,转化至JM109感受态细胞进行蓝白斑筛选。最后提取质粒进行电泳检测、PCR鉴定并将带有目的基因的质粒送至上海生工进行测序,测序结果与预期序列相符,测序正确的重组质粒命名为pUCm-T-AusfaeA。Blast分析结果表明该序列与 Aspergillus niger faeA(Y09330.2)、Aspergillusflavus NRRL3357 faeA(XM_002380132.1)及 Aspergillus oryzae RIB40 faeA(XM_001818700.1)的序列同源性分别为99%、70.9%和70.6%。
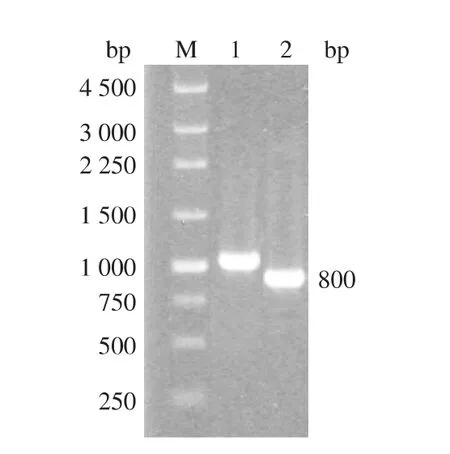
图1 AusfaeAPCR产物电泳图Fig.1 Electrophoretic profile of PCR products of AusfaeA
2.2 pPIC9K-AusfaeA的构建与鉴定
按照1.7中的方法,将pUCm-T-AusfaeA和pPIC9K质粒分别用EcoR I和Not I进行双酶切,酶切后分别进行电泳检测。前者得到了两条条带,一条在2.7 kb左右,另一条位于800 bp左右;后者得到一条位于9 kb左右的条带。将800 bp和9 kb左右的条带割胶回收。然后将回收的产物连接过夜,转化DH5α感受态细胞。用PCR检测的方法筛选目的重组子,将目的重组子送至上海生工进行测序,结果表明基因没有发生突变,且读码框完全正确。
2.3 重组GS115的构建及高拷贝重组子的筛选与表达
按照1.8中的方法得到高拷贝的GS115/AusfaeA和GS115/9K重组酵母,提取酵母基因组利用通用引物5'-AOX和3'-AOX做PCR检测,如图2所示,因为P.pastoris基因组中本身含有一个醇氧化酶基因(AOX1),其大小为2.1 kb,而pPIC9k中包含醇氧化酶-1(AOX1)基因的启动子和转录终止子(5'AOX1 和 3'AOX1),它们被多克隆位点(MCS)分开,外源基因可以在此插入,所以当利用通用引物5'-AOX和3'-AOX进行PCR鉴定时,重组GS115均出现两条条带,GS115/AusfaeA的PCR产物分别位于2.1 kb和1.3 kb附近,而GS115/9K的PCR产物分别位于2.1 kb和0.5 kb附近,结果表明AusfaeA已经成功整合入毕赤酵母GS115基因组内。然后,随机挑取鉴定过的重组酵母菌株进行诱导表达,8 000 r/min离心5 min收集表达产物备测。
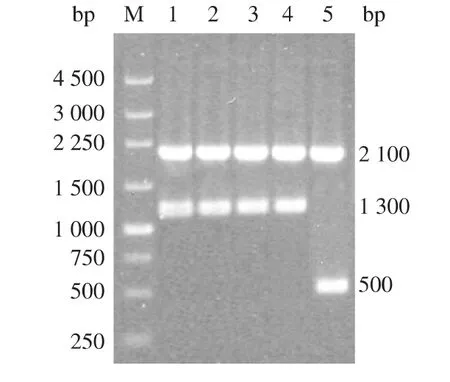
图2 毕赤酵母重组子基因组PCR验证Fig.2 Verification of P.pastoris transformants by PCR
2.4 reAusFaeA的酶活及SDS-PAGE分析
将表达产物按1.9的方法测定reAusFaeA的酶活,结果表明GS115/AusfaeA发酵上清液中reAusFaeA的酶活可达到7.05 U/mL,而GS115/9K的发酵上清液中并未检测到阿魏酸酯酶活性。经纯化后的reAusFaeA比酶活为29.4 U/mg。SDS-PAGE结果显示,GS115/AusfaeA的表达产物在约36 000处可见明显的特异性目的蛋白条带 (图3泳道2-3),而GS115/9K的表达产物在该处并无特异性条带(图3泳道1)。结果表明reAusFaeA的表观分子量高于理论计算相对分子质量(28 300)。这可能由于reAusFaeA在P.pastoris表达过程中发生了糖基化或磷酸化等翻译后修饰。用软件NetNGlyc 1.0(http://www.cbs.dtu.dk/services/NetNGlyc/) 和 NetOGlyc 3.1 (http://www.cbs.dtu.dk/services/NetOGlyc/) 分 别对N和O糖基化位点进行预测。结果表明AusFaeA氨基酸序列中含有1个N-糖基化位点。
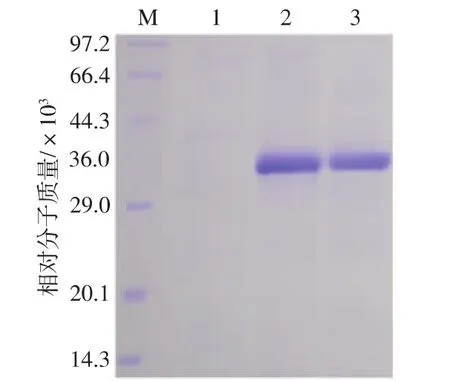
图3 reAusFaeA的SDS-PAGE分析Fig.3 SDS-PAGE analysis of reAusFaeA
2.5 reAusFaeA的酶学鉴定
按照1.10.1中的方法测定reAusFaeA的最适温度及温度稳定性。结果表明,该酶的最适反应温度为45℃,在40~50℃的范围内催化活性较高 (图4);由图5可以看出,该酶在45℃的条件下相当稳定,处理60 min残余酶活仍能达到89.3%;当温度高于45℃时,残余酶活下降较快,在55℃下处理20 min后残余酶活性仅为7.91%。这也说明该酶的热稳定性有待进一步地提高。
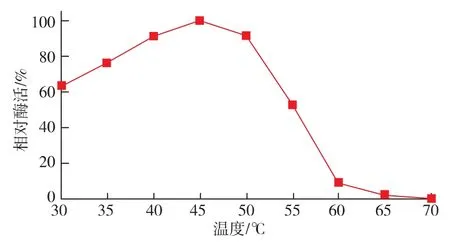
图4 温度对reAusFaeA酶活力的影响Fig.4 Effect of temperature on the activity of the reAusFaeA
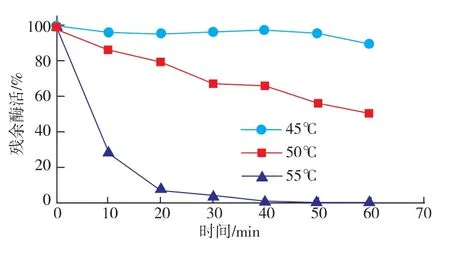
图5 reAusFaeA的温度稳定性Fig.5 Effect of temperature on enzyme stability
按照1.10.2中的方法测定reAusFaeA的最适pH及pH稳定性。结果表明,该酶的最适pH为5.0,当pH低于3.5和高于7.5时该酶几乎没有催化活性(图6)。由图7可知,该酶在pH 4.0~6.0的范围内处理1 h后残余酶活仍能达到60%以上;当pH值低于4.0和高于6.5时该酶的稳定性较差。
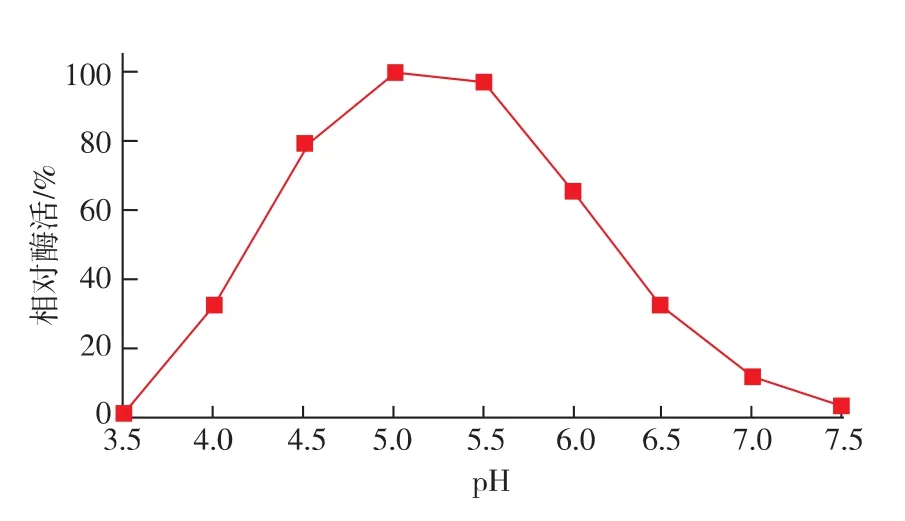
图6 pH对reAusFaeA酶活力的影响Fig.6 Effect of pH on the activity of the reAusFaeA
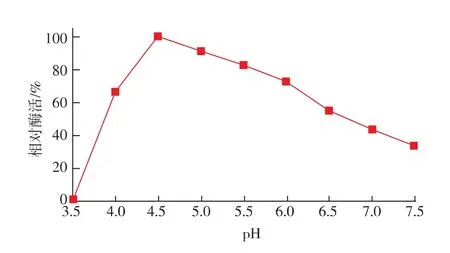
图7 reAusFaeA的pH稳定性Fig.7 Effect of pH on enzyme stability
按照1.10.3中的方法,研究了不同金属离子及EDTA对reAusFaeA酶活性的影响。结果表明,大多数被测的金属离子及EDTA对该酶的活性影响不大,说明该酶具有被广泛应用的潜力。
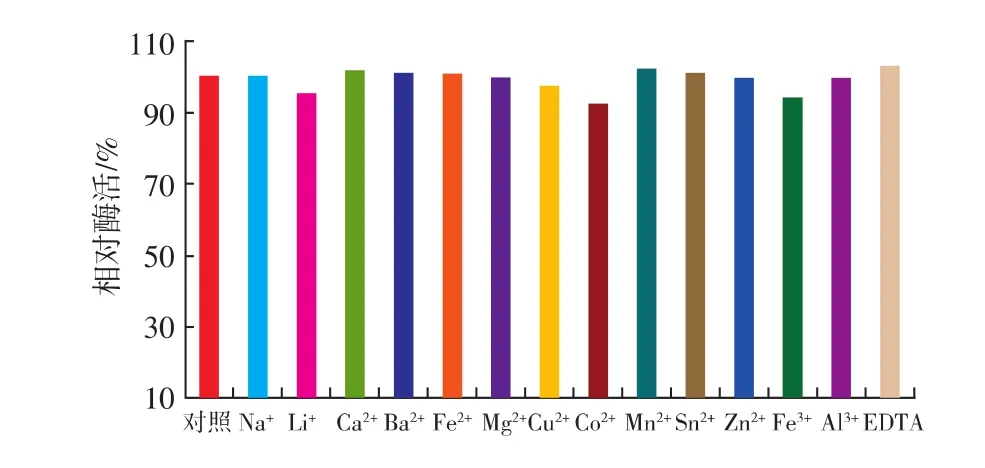
图8 金属离子和EDTA对AusFaeA酶活力的影响Fig.8 Effects of various metal ions and EDTA on the activity of the reAusFaeA
3 结语
阿魏酸酯酶有着重要的工业应用价值,通过基因工程技术实现阿魏酸酯酶的异源高效表达是降低其工业应用成本、适应工业应用条件的有效手段。国内对于阿魏酸酯酶异源表达的相关报道较少。Mathew等[16]对Aspergillus flavipes进行了液体深层发酵获取了阿魏酸酯酶,以阿魏酸甲酯为底物,最高酶活可达6.82 U/mL。
作者首次成功实现了AusfaeA在P.pastoris GS115中的分泌表达,毕赤酵母是一个优良的表达体系,其具有操作简便、可高水平地胞外表达、可进行糖基化、二硫键形成等真核翻译后修饰等优点[17]。以阿魏酸甲酯为底物,reAusFaeA粗酶液的酶活可达到7.05 U/mL,经纯化后,该酶的比酶活为29.4 U/mg;SDS-PAGE表明出reAusFaeA的表观分子量约为36.0 kDa;初步的酶学性质研究表明该酶的最适反应温度为45℃,并在45℃下相当稳定;最适反应pH为5.0,在pH 4.0~6.0的条件下处理1 h仍有60%以上的残余酶活;EDTA和大多数被测金属离子对该酶的活性影响不大;这些优良的酶学特性表明了该酶具有被广泛应用的潜力。遗憾的是,该酶在55℃的条件下处理20 min后残余酶活性仅为7.91%,表明该酶的热稳定性较差。若要将其与某些耐热的木聚糖酶共同使用,今后还需对其热稳定性进行必要的理性改造。
[1]许晖,孙兰萍,张斌,等.微生物阿魏酸酯酶的研究进展[J].中国酿造,2008,10:11-15.XU Hui,SUN Lan-ping,ZHANG Bin,et al.Advance research on feruloyl esterases produced by microbes[J].China Brewing,2008,10:11-15.(in Chinese)
[2]Vries RP.DE,Visser J.Aspergillus enzymes involved in degradation of plant cell wall polysaccharides[J].Microbiology and Molecular Biology Reviews,2001,65(4):497-522.
[3]Luonteri E,Kroon P A,Tenkanen M,et al.Activity of an Aspergillus terreus α-arabinofuranosidase on phenolic-substituted oligosaccharides[J].Journal of Biotechnology,1999,67:41-48.
[4]Kroon PA,Garcia-Conesa MT,Fillingham IJ,et al.Release of ferulic acid dehydrodimers from plant cell walls by feruloyl esterases[J].Journal of the Science of Food and Agriculture,1999,79:428-434.
[5]Crepin V F,Faulds C B,Connerton I F.Functional classification of the microbial feruloyl esterases[J].Appl Microbiol Biotechnol,2004,63:647-652.
[6]Williamson G J,Faulds C B.The purification and characterization of 4-hydroxy-3-methoxycinnamic (ferulic) acid esterase from Streptomyces olivochromogenes[J].Joumal of General Microbiology,1991,137:2339-2345.
[7]胡雪松,李夏兰.阿魏酸酯酶基因克隆表达调控及其应用进展[J].生物技术通报,2009,12:11-16.HU Xue-song,LI Xia-lan.Review on cloning,expression,regulation and applicationsof feruloyl esterase genes[J].Biotechnology Bulletin,2009,12:11-16.(in Chinese)
[8]刘晶晶,曹阳春,杨红建.阿魏酸酯酶表达体系与酶蛋白拓扑空间结构分析[J].中国畜牧兽医,2009,36(12):26-30.LIU Jing-jing,CAO Yang-chun,YANG Hong-jian.Ferulic acid esterase expression system and enzyme protein topology analysis of the spatial structure[J].China Animal Husbandry And Veterinary Medicine,2009,36(12):26-30.(in Chinese)
[9]Nethaji B M,Pattabhi V.Structure of 3-(4-hydroxy-3-rnethoxyphenyl)-2-propenoic acid (ferulic acid).ActaCryst,1988,C44:275-277.
[10]刘子立,欧仕益.离子交换法纯化阿魏酸的研究[J].广州食品工业科技,2003,20:44-47.LIU Zi-li,OU Shi-yi.Extraction of ferulic acid from enzymatic hydrolysate by using ion-exchange adsorption[J].Guangzhou Food Science and Technology,2003,20:44-47.(in Chinese)
[11]李夏兰,程珊影,杨道秀,等.阿魏酸酯酶和木聚糖酶协同降解麦糟[J].化工进展,2012,31(5):1096-1102.LI Xia-lan,CHEN Shan-ying,YANG Dao-xiu,et al.Utilization of feruloyl esterase and xylanase for the degradation of brewers’spent grain[J].Chemical Industry And Engineering Progress,2012,31(5):1096-1102.(in Chinese)
[12]Li J F,Zhao S G,Tang C D,et al.Cloning and functional expression of an acidophilic beta-mannanase gene (Anman5A) from Aspergillus niger LW-1 in Pichia pastoris[J].Journal Of Agricultural And Food Chemistry,2012,60(3):765-773.
[13]Tang C D,Guo J,Wu M C,et al.Cloning and bioinformatics analysis of a novel acidophilic β-mannanase gene,Auman5A,from Aspergillus usamii YL-01-78[J].World Journal of Microbiology and Biotechnology,2011,27(12):2921-2929.
[14]Bradford M.A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dyebinding[J].Analytical Biochemistry,1976,72:248-254.
[15]Zhang S B,Pei X Q,Wu Z L.Multiple amino acid substitutions significantly improve the thermostability of feruloyl esterase A from Aspergillus niger[J].Bioresource Technology,2012,117:140-147.
[16]Mathew S,Abraham T E.Studies on the production of feruloyl esterase from cereal brans and sugar cane bagasse by microbial fermentation[J].Enzyme and Microbial Technology,2005,36:565-570.
[17]谌斌,唐雪明,沈微,等.粗糙脉孢菌漆酶基因的克隆及在毕赤酵母中的初步表达[J].食品与生物技术学报,2012,23(4):43-47.CHEN Bin,TANG Xue-ming,SHEN Wei,et al.Cloning of a laccase gene from Neurospora crassa and it’s preliminary expression in Pichia pastoris[J].Journal of Food Science and Biotechnology,2012,23(4):43-47.(in Chinese)
