西北太平洋柔鱼资源丰度的空间尺度分析
2013-02-15杨铭霞陈新军冯永玖
杨铭霞,陈新军、2、3,冯永玖、2、3
(1.上海海洋大学 海洋科学学院,上海201306;2.上海海洋大学 大洋生物资源开发和利用上海市高校重点实验室,上海201306;3.上海海洋大学 大洋渔业资源可持续开发省部共建教育部重点实验室,上海201306)
西北太平洋柔鱼Ommastrephes bartramii是一年生的短周期大洋性洄游种类[1-2],是中国鱿钓船的主要捕捞对象。柔鱼渔场的分布与黑潮、亲潮势力强弱及其分布密切相关,柔鱼在夏秋季北上期间,一般分布在表层水温较高的黑潮前锋附近以及等温线分布密集的暖冷水交汇区[3]。柔鱼常常以群体的形式,分布在某种特定的空间或时间上,它们具有高度的空间和时间异质性[4]。
传统统计学通常把研究对象作为独立存在的随机变量,不考虑与影响因素之间的空间相关性,因此存在很大的局限性。而地统计学避免了经典统计学中忽视空间位置和方向的缺陷,提供了一个有效地分析和解释空间数据的方法[5]。地统计学以变异函数为工具,研究那些在空间分布上既具有结构性又有随机性,或有空间相关和依赖性的自然现象[6-8]。而尺度问题是所有生态学研究的基础,正逐渐被越来越多的生态学家认识[9]。本研究中主要采用地统计学方法对西北太平洋柔鱼资源丰度空间分布的尺度进行探讨,并结合柔鱼群体自身的种群结构特征以及西北太平洋的海洋环境因素,分析引起这些差异的原因,旨在为更好地研究西北太平洋柔鱼资源丰度的空间分布提供参考。
1 材料与方法
1.1 材料
2011年西北太平洋柔鱼生产统计数据来自上海海洋大学鱿钓技术组,其内容主要包括作业时间、作业海区经纬度以及单船产量等。
1.2 方法
1.2.1 数 据 处 理 取 样 范 围 为150° ~160°E、38° ~48°N,以西北太平洋柔鱼盛渔期的8月份为例进行研究。将8月柔鱼单船日产量(CPUE)数据分别按经纬度10'×10'、20'×20'、30'×30'、40'×40'、50'×50'、60'×60'、70'×70'、80'×80'、90'×90'、100'×100'、110'×110'和120'×120'以上等12 个空间尺度进行划分,形成不同大小的栅格数据(图1),对每一统计单位的站点数据取算术平均值。
1.2.2 地统计学方法 本研究中借助ArcGis 9.0地统计模块对数据进行处理分析。对于数据序列Z(xi),i=1,2,…,n,样本的半变异函数r(h)用下式计算:
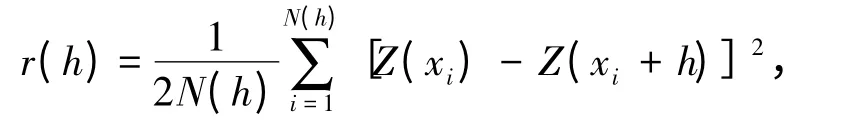
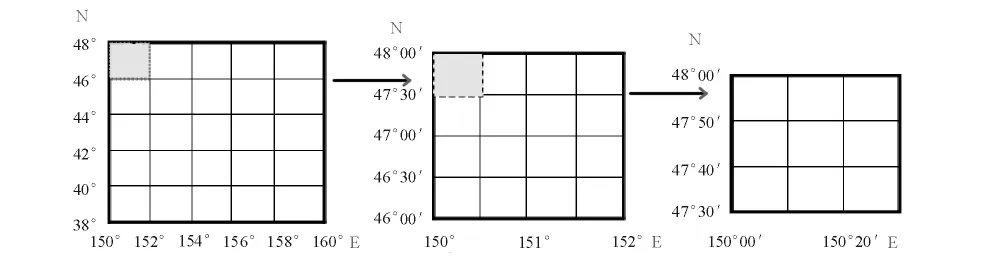
图1 不同尺度下取样站点数据分布示意图Fig.1 The illustration of sampling data in different spatial scales
式中:N(h)为被h 分割的数据对(xi,xi+h)的对数;Z(xi)和Z(xi+h)分别为xi和xi+h 处样本的测量值;h 为分隔两样点的距离(小于最大间距)。半变异函数曲线通常用球状模型、指数模型和高斯模型进行拟合,筛选出最优模型(图2)。其中C0为块金值,其大小反映变量的随机程度;C+C0为基台值,其大小反映变量变化幅度的大小,C 为偏基台值;C0/(C+C0)的值可用来判断系统内变量的自相关程度,<25%时为强空间自相关性,25% ~75%时为中等自相关性,>75%为弱自相关性[10];a 为变程,表示以a 为半径的领域内任何其他Z(xi)和Z(xi+h)间存在的空间自相关性,或者Z(xi)和Z(xi+h)相互有影响。
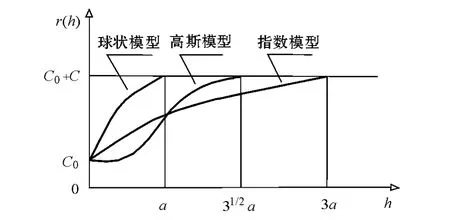
图2 球状模型、指数模型和高斯模型示意图Fig.2 Diagrams of Spherical model,Exponential model and Gaussian model
2 结果与分析
2.1 柔鱼CPUE 的空间分布特征
8月柔鱼作业区域主要集中在154° ~158°E、39° ~44°N,累计作业次数达到3 360次,累计总产量为9 062.56 t。作业渔船分布极为密集(图3)。8月份CPUE 为1 ~10 t/d,约占总作业次数的77.38%;CPUE 达10 t/d 以上的作业次数约占总数的1.87%;总平均CPUE 为2.70 t/d。
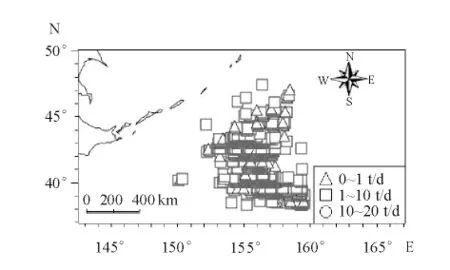
图3 西北太平洋柔鱼CPUE 空间分布图Fig.3 Spatial distribution of CPUE for flying squid Ommastrephes bartramii in the northwest Pacific Ocean
2.2 相关变量分析
随着分布尺度的递增,最优模型的选择以及各个参数在某种程度上显现出部分规律性(表1)。从表1可见:当尺度≥80'×80'时,均以高斯模型拟合的最好;从方向角分布来看,当尺度<80'×80'时,随着尺度的增大,方向角从WN-ES 方向逐渐向N-S 偏转,当尺度>80'×80'时则呈现无规律性,说明随机性比较大。同样,从各尺度下的变程趋势也不难发现(图4),随着尺度的递增,变程逐渐增大,而当尺度>80'×80'时则无又规律可言。
从图5可见:块金值与基台值的走势基本保持一致,当尺度为40'×40'、60'×60'、90'×90'时分别出现了高峰,说明在这3 个尺度下块金值与基台值变化幅度相对较大,由随机性引起的可能性也较大。根据以上结果可初步推断,当尺度>80'×80'时,变量由随机性产生的可能性过大,可能不太适合用来研究西北太平洋柔鱼的空间异质性。

表1 西北太平洋柔鱼各尺度下有关参数Tab.1 Parameters for various scales in flying squid Ommastrephes bartramii in the northwest Pacific Ocean

图4 西北太平洋柔鱼变程随尺度的变化Fig.4 Variation trend of ranges for each scale in flying squid O.bartramii in the Northwest Pacific Ocean
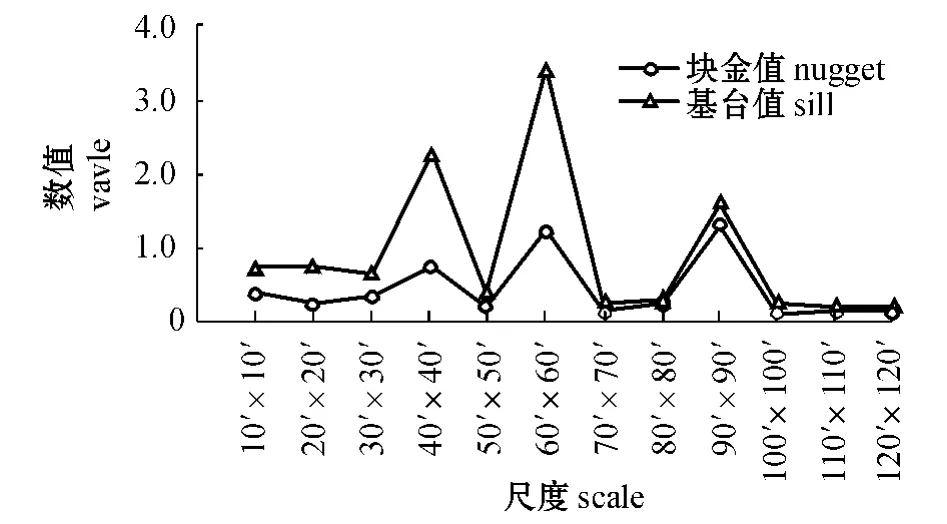
图5 西北太平洋柔鱼块金值和基台值随尺度的变化Fig.5 Values of Nugget and Sill for various scales in flying squid O.bartramii in the Northwest Pacific Ocean
2.3 空间自相关程度
从图6可见:空间的自相关性大体上处于中等水平,在各尺度下不存在强空间自相关性。其中当经纬度尺度为80'×80'和90'×90'时,呈现弱空间自相关性;当经纬度尺度为40'×40'时,空间自相关性最强,达69.4%(1-块金值/基台值)。

图6 西北太平洋柔鱼块金值与基台值之比随尺度的变化Fig.6 Ratio of Nugget to Sill for various scales in flying squid O.bartramii in the northwest Pacific Ocean
3 讨论
3.1 渔场分布及其与环境的关系
本研究中只采用了2011年8月的渔获量数据,从该月CPUE 空间分布来看,作业船只主要集中在154° ~158°E、39° ~44°N 海域,该海域是中国鱿钓渔船传统的重要作业渔场。8月产量占全年总渔获量的28.8%,平均CPUE 为2.70 t/d,高于全年平均CPUE(1.93 t/d)。8月黑潮暖流与亲潮寒流相遇,受黑潮强势力影响,在154° ~158°E、39° ~44°N 海域形成流隔渔场。在150° ~160°E 海域,表温为13 ~21 ℃,总体上与往年同期适宜表温范围基本一致[11-15]。柔鱼高密度分布海域的表温为18 ~20 ℃,与1998年、2003—2006年等暖水年相比[16-19],最适表温偏高1 ℃左右;与2001年、2009年等黑潮大弯曲及1996年、2000年、2001年亲潮势力强劲的冷水年相比[20-21],最低适宜温度高出3 ~5 ℃。8—10月为盛渔期,由于黑潮势力较强,渔汛延长,产量较高。
3.2 多尺度效应
为了提高海洋渔业数据的检索和操作效率,本研究中将庞大的数据量进行网格化[22],划分为10'×10'、20'×20'、…、120'×120'共12 个等级的空间分辨率。将10'×10'、20'×20'、30'×30'视为小尺度,40'×40' ~70'×70'视为中尺度,80'×80'以上视为大尺度。根据鲁学军等[23]提出的“空间分辨率圆锥”,在各分辨率下反映区域地理系统内部组织构造单元之间包含与被包含的空间关系将不同,所发生的地理事件的规模和性质也将不同。
柔鱼群体地理空间分布的研究,可反映种群内部结构机理。小尺度空间分辨率下,块金值和基台值的变化幅度保持相对稳定,说明小空间分辨率内柔鱼群体的内部结构相对稳定,没有太大差异。这反映了柔鱼在索饵期间群体分布的空间格局为聚集型,这种现象在张寒野等[24]的研究结果中也有发生。研究其空间分布格局或许能解释柔鱼在各生长时期内不同的行为表现,如为了减轻生长空间的压力和降低种内竞争,柔鱼进行大范围洄游以及多季节产卵等[25]。
柔鱼的生长除了与其自身群体结构及其同类有关之外,还受到周围海洋环境的影响。中尺度空间分辨率下,块金值和基台值出现了大幅度的波动,而变程的变化率保持稳定,这说明在一定的空间分辨率下柔鱼群体受到某种特定环境因素的影响,随着分辨率的递增,空间自相关性的半径范围呈线性增大,但不影响其空间自相关性。据日本学者对柔鱼冬-春生群体和秋生群体洄游模式的推定[26-27],本研究中8月西北太平洋柔鱼很有可能为冬-春生群体,此群体从产卵场洄游至索饵场,也是黑潮与亲潮的交汇区,此间柔鱼的主要移动路线与黑潮暖水系以及亲潮分支关系密切。
随着空间尺度的增大,块金值和基台值在90'×90'尺度下出现一个高峰,变量的随机性及变化幅度较大,之后则趋于稳定,可能进入了另一个地理事件。但从空间相关性来看,大尺度空间分辨率下,变程上下波动,空间自相关性范围时大时小,从某种程度上说明了柔鱼群体的空间分布受多因素的交互影响。而由图6可见,大尺度下柔鱼群体的空间自相关性明显比中小尺度的弱。这可能是因为大范围内柔鱼群体受海洋大环境的影响,因而减弱了其自身结构特征。
陈新军等[28]对西北太平洋150° ~165°E 海域柔鱼日产量与时间、空间、表温、表温距平均值等环境因子进行了灰色关联分析,认为8—9月在38° ~44°N 海域100 m 水层内形成明显的温跃层,200 m 以下水温几乎无变化,夜间柔鱼主要在温跃层附近或者在温跃层与海面之间游泳,与柔鱼空间自相关体现的聚集性刚好相吻合,因此温跃层也是影响柔鱼空间分布的一个重要环境因子。柔鱼具有昼夜垂直移动习性,在索饵海域夜间主要游泳层在水深40 m 以上,而白天主要游泳层为水深150 ~300 m[3]。由于受到数据资料的限制,没有将夜间和白天捕获的柔鱼分开,不能获得不同水层的数据资料,因此,对柔鱼资源三维空间格局分布的研究工作受到了限制。
本研究中借鉴了地统计学方法,引入了空间尺度分析的思想,探讨了2011年8月西北太平洋柔鱼群体的空间分布以及自相关性特征。初步分析了在各个尺度下柔鱼资源分布产生的效应,并结合柔鱼群体自身结构特点及海洋环境因素进行了解释。对柔鱼资源的不同尺度空间格局进行研究,既可减少单一尺度下对研究结果造成的误差,又可减少用大尺度研究时忽略局部环境特点或用小尺度研究时脱离了大环境背景所造成的误差。在今后的研究中,可考虑用三维时空分布格局和专家模型[29]来研究西北太平洋柔鱼群体的空间分布特征。
[1]Yatsu A,Tanaka H,Mori J.Population structure of the neon flying squid,Ommastrephes bartamii,in the North Pacific[C]//Okutani T.Contributed papers to International Symposium on Large Pelagic Squids.Tokyo:Japan Marine Fishery Resources Research Center,1998:31-48.
[2]Nagasawa K,Mori J,Okamura H.Parasites as biological tags of stocks of neon flying squid(Ommastrephes bartramii)in the North Pacific Ocean[C]//Okutani T.Contributed Papers to International Symposium on Large Pelagic Squids.Tokyo:Japan Marine Fishery Resources Research Center,1998:49-64.
[3]王尧耕,陈新军.世界大洋性经济柔鱼资源及其渔业[M].北京:海洋出版社,2005:138-150.
[4]王政权.地统计学及在生态学中的应用[M].北京:科学出版社,1999:1-4.
[5]侯景儒,黄竞先.地质统计学的理论与方法[M].北京:地质出版社,1990.
[6]刘丹,姜勇,梁文举,等.沈阳市郊耕地土壤交换性Ca 含量的空间异质性特征[J].生态学杂志,2003,22(4):6-9.
[7]Isaaks E H,Srivastava R M.An introduction to applied geostatistics[M].NewYork:Oxford University Press,1989.
[8]Webster R.Quantitative spatial analysis of soil in the field[J].Adv Soil Sci,1985,3:1-70.
[9]张娜.生态学中的尺度问题:内涵与分析方法[J].生态学报,2006,26(7):2340-2355.
[10]刘杏梅.基于GIS和地统计学的不同尺度水稻田土壤养分时空变异及其机理研究[D].杭州:浙江大学,2010.
[11]陈新军,刘必林.2004年北太平洋柔鱼钓产量分析及作业渔场与表温的关系[J].湛江海洋大学学报,2005,25(6):41-45.
[12]陆化杰,陈新军.2006年北太平洋柔鱼作业渔场时空变化及其与表温的关系[J].广东海洋大学学报,2008,28(1):93-97.
[13]陈新军,田思泉,许柳雄.西北太平洋海域柔鱼产卵场和作业渔场的水温年间比较及其与资源丰度的关系[J].上海水产大学学报,2005,14(2):168-175.
[14]沈新强,樊伟,崔雪森.西北太平洋柔鱼渔场分布与水温关系的研究[J].海洋水产研究,2004,25(3):10-14.
[15]陈新军,田思泉.西北太平洋海域柔鱼的产量分布及作业渔场与表温的关系研究[J].中国海洋大学学报,2005,35(1):101-107.
[16]陈新军,许柳雄,田思泉.北太平洋柔鱼资源与渔场的时空分析[J].水产学报,2003,27(4):334-342.
[17]陈峰,陈新军,钱卫国,等.水温变动对2009年西北太平洋柔鱼产量下降的影响[J].广东海洋大学学报,2010,30(1):65-71.
[18]陈峰,陈新军,刘必林,等.西北太平洋柔鱼渔场与水温垂直结构关系[J].上海海洋大学学报,2010,19(4):495-504.
[19]范江涛,陈新军,曹杰,等.西北太平洋柔鱼渔场变化与黑潮的关系[J].上海海洋大学学报,2010,19(3):378-384.
[20]唐峰华,崔雪森,樊伟,等.北太平洋柔鱼渔获量与海洋环境关系的遥感学研究[J].海洋技术,2011,30(2):7-12.
[21]邵全琴,马巍巍,陈卓奇,等.西北太平洋黑潮路径变化与柔鱼CPUE 的关系研究[J].海洋与湖沼,2005,36(2):111-122.
[22]季民,靳奉祥,李云岭,等.海洋渔业专题属性数据多尺度综合与表达[J].测绘通报,2004(6):28-31.
[23]鲁学军,励惠国,陈述彭.地理时空等级组织体系初步研究[J].地球信息科学,2000,2(1):60-66.
[24]张寒野,胡芬.冬季东海太平洋褶柔鱼的空间异质性特征[J].生态学杂志,2005,24(11):1299-1302.
[25]杨林林,姜亚洲,程家骅.东海太平洋褶柔鱼生殖群体的空间分布及 其 与 环 境 因 子 的 关 系[J].生 态 学 报,2010,30(7):1825-1833.
[26]村田守.北太平洋におけるいか流し网漁場の海洋環境及びアカイカの分布洄游[R].北海道:北海道区水産研究所(第17 号),1990:144-148.
[27]谷津明彦.北太平洋における釣り調査によるアカイカの分布(1976-1983年)[R].清水:遠洋水産研究所研究報告(29),1992:13-37.
[28]陈新军,许柳雄.北太平洋150°E ~165°E 海域柔鱼渔场与表温及水温垂直结构的关系[J].海洋湖沼通报,2004(2):36-44.
[29]于红,刘溪婧.基于知识库的渔业领域本体学习算法[J].大连海洋大学学报,2011,26(2):168-172.
