高温对虾夷扇贝存活率、耗氧率和排氨率的影响
2013-02-15郝振林丁君贲月常亚青
郝振林,丁君,贲月,常亚青
(大连海洋大学 农业部北方海水增养殖重点实验室,辽宁大连116023)
虾夷扇贝Mizuhopecten yessoensis是大型冷水性双壳贝类,主要分布于俄罗斯远东海、千岛群岛,日本北海道、本洲北部,以及朝鲜北部海域[1]。20世纪80年代初,中国从日本引进虾夷扇贝,进行筏式网笼养殖和底播增殖。1990年虾夷扇贝筏式养殖形成较大规模,主要养殖区集中在长海县的海洋岛、獐子岛和小长山海域[2]。近年来,养殖虾夷扇贝出现大批死亡现象,特别是1 龄贝、15月龄至20月龄的虾夷扇贝,每年7—8月份死亡率明显增加。2009年,长海县虾夷扇贝出现大面积死亡,3—4月份筏式养殖的1 龄贝和2 龄贝死亡率约50%;7月份后,死亡率高达80%,给养殖者带来了巨大的经济损失[3]。虾夷扇贝夏季出现大规模死亡的原因目前尚在进一步探讨中,有学者认为虾夷扇贝的高死亡率可能与海水温度过高有关[4]。高温是水产动物在养殖过程中面临的一个重要威胁,但关于高温对虾夷扇贝影响的研究鲜见报道。为此,作者以獐子岛底播贝虾夷扇贝为试验材料,在实验室内设计了两个试验,以检测虾夷扇贝对温度突变的耐受能力以及在不同高温水平下的存活、行为和生理反应,旨在为后期进行耐高温型虾夷扇贝新品系的培育及健康养殖提供参考。
1 材料与方法
1.1 材料
试验用虾夷扇贝为獐子岛底播贝,保湿状态下运到大连海洋大学,清理干净后,暂养于农业部北方海水增养殖重点实验室可控温独立循环品字形水槽内。暂养期间,海水温度为(15±1)℃,盐度为30 左右,每天定量投喂螺旋藻粉和鼠尾藻粉,每2 d 全量换水1次。驯养7 d 后,挑选活力较强的健康个体用于试验。
1.2 方法
1.2.1 试验温度梯度的设定 试验设15(对照)、2 0、2 2、2 4、2 6 ℃5 个温度梯度。试验Ⅰ:将在15 ℃下暂养的虾夷扇贝分别直接放到20、22、24、26 ℃的海水中,并在1、2、4、8、12、24、48、96 h 时测定虾夷扇贝的存活率。试验Ⅱ:将在15℃下暂养的虾夷扇贝分别驯化到20、22、24、26℃,升温幅度为1 ℃/d,饲养7 d 后观察虾夷扇贝的行为特征,测定其存活率、耗氧率和排氨率。
每个盐度梯度设3 个平行。测定耗氧率和排氨率所用容器为4 L 自封袋,每个自封袋中放1 个扇贝。用虹吸法采水并测定耗氧率和排氨率,虹吸前用手轻轻揉捏自封袋,使各处溶氧及氨氮保持均衡,试验时间持续2 h。
1.2.2 试验指标的测定

其中:DO0和DOt分别为试验开始和结束时水体中的DO 含量(mg/L);N0和Nt分别为试验开始和结束时水体中的(μmol/L);V 为试验用水体积(L);W 为试验贝软组织干质量(g);t 为试验时间(h)。
3)温度对虾夷扇贝代谢率的影响强度(Q10)。计算公式为
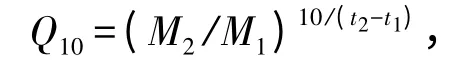
其中:M1和M2分别为试验开始(t1)和试验结束(t2)时扇贝的代谢率。
4)氧氮比(O∶N)。用呼吸氧原子数与排出氨态氮原子数之比来计算。
试验结束后,测定虾夷扇贝的壳长(mm)和壳高(mm),称量湿体质量和软体部干质量(g)(60 ℃下烘干至恒重)。
1.3 数据处理
采用SPSS 10.0 软件进行单因子方差分析(One-Way ANOVA)、组间多重比较(LSD ANOVA)和相关回归分析,显著性水平设为0.05。
2 结果与分析
2.1 生物学指标
经测定,试验用虾夷扇贝的壳长为(95.38±6.12)mm,壳高为(96.12±7.03)mm,湿质量为(107.38±2.21)g,软体部干质量为(4.67±0.57)g。
2.2 存活
温度突变时虾夷扇贝的存活率如图1所示。从图1可见:经96 h 胁迫,15 ~24 ℃处理组虾夷扇贝的存活率均高于82.29%,且组间无显著性差异(P>0.05);但26 ℃处理组虾夷扇贝在不同胁迫时间下的存活率却存在显著性差异(P<0.05),经4 h 胁迫时,其存活率为87.50%,经8 h 胁迫时,存活率为62.50%,经12 h 胁迫时,存活率为0。虾夷扇贝在8、12、24、48、96 h 时的半致死温度(LT50)分 别 为 27.52、24.41、24.37、24.24、23.81 ℃。
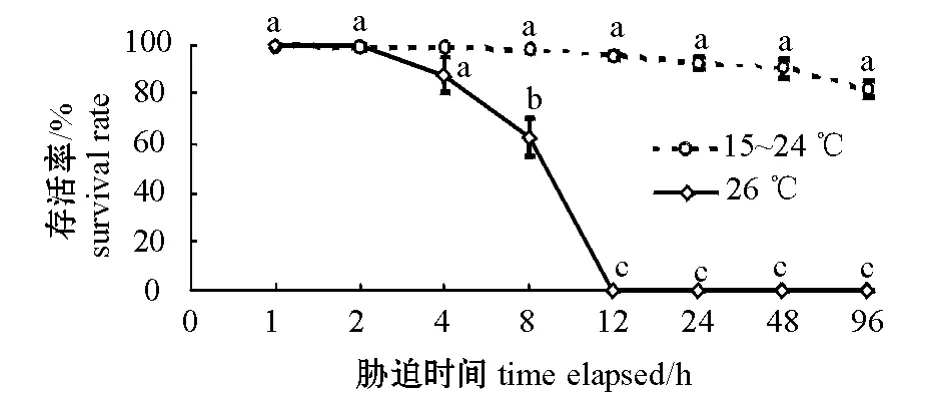
图1 温度突变对虾夷扇贝存活率的影响Fig.1 The changes in survival rate of yesso scallop Mizuhopecten yessoensis exposed to different abrupt temperature changes
从图2可见:在试验温度范围内,虾夷扇贝的存活率随温度的变化呈逐渐下降的趋势,其中15℃处理组存活率最高(100%),26 ℃处理组虾夷扇贝的存活率最低(26.13%);15 ~22 ℃处理组存活率均高于85%,且组间无显著性差异(P >0.05),但均显著高于24、26 ℃组(P<0.05)。拟合得到存活率(S)与温度(t)之间的相关方程为S=-0.0543t2+0.1397t+0.908(R2=0.9868,n=75)。
2.3 行为特征
虾夷扇贝在不同温度处理组中的行为特征有明显差别。在15 ℃处理组中,虾夷扇贝双壳始终处于微闭状态(双壳腹缘中点的直线距离为5 ~8 mm),在受到惊吓时,其双壳立即紧闭,闭合力较大;在20 ~24 ℃处理组中,虾夷扇贝双壳开口较1 5℃处理组略大(双壳腹缘中点的直线距离为8 ~16 mm),受到惊吓时,双壳立即紧闭,闭合力较大;在26 ℃处理组中,虾夷扇贝双壳开口非常大(双壳腹缘中点的直线距离为20 ~35 mm),受到惊吓时,双壳不能紧闭,即使其外套膜受到手指刺激时,双壳亦不能紧闭,且闭合力很弱。

图2 温度缓变对虾夷扇贝存活率的影响Fig.2 The changes in survival rate of yesso scallop M.yessoensis exposed to different slow temperature changes
2.4 耗氧率
从图3可见:虾夷扇贝的耗氧率随温度的升高呈先升高后降低的趋势,其中22 ℃处理组虾夷扇贝的耗氧率最高[1.56 mg/(g·h-1)],15 ℃处理组的耗氧率最低[0.60 mg/(g·h-1)],20 ~24 ℃处理组间耗氧率无显著性差异(P >0.05),但均显著高于15、26 ℃处理组(P<0.05)。拟合得到温度(t)与耗氧率(RO)之间的关系式为
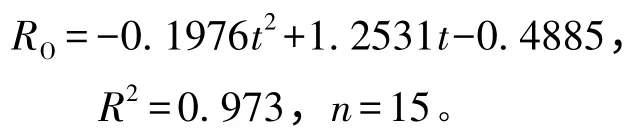
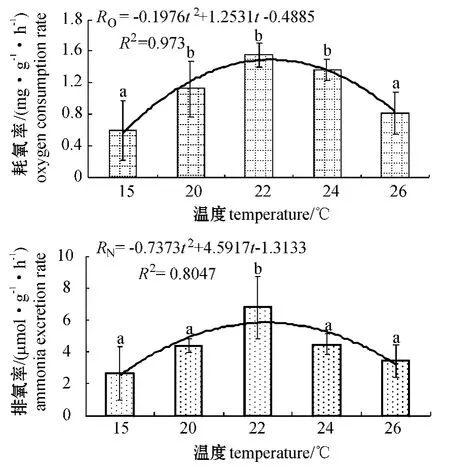
图3 不同温度下虾夷扇贝的耗氧率和排氨率Fig.3 Oxygen consumption rates,and ammonia-N excretion rates in yesso scallop M.yessoensis at different temperature
2.5 排氨率
从图3可见:虾夷扇贝排氨率的变化趋势同耗氧率相似,其中,22 ℃处理组虾夷扇贝的排氨率最高[6.81 μmol/(g·h)],且显著高于其他各温度组(P<0.05),而其余组间均无显著性差异(P>0.05)。拟合得到温度(t)与排氨率(RN)之间的关系式为
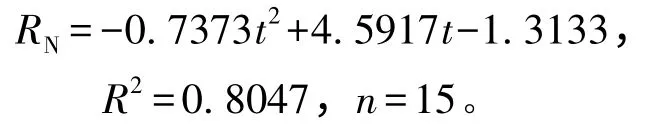
2.6 代谢水平
从图4可见:O/N 值随温度的升高呈先增大后下降的趋势,整个试验过程中,O/N 值为14.92 ~20.79,其中在24 ℃时O/N 值达到最大(20.79),且显著高于其他各温度组(P<0.05),而其余组间均无显著性差异(P>0.05)。
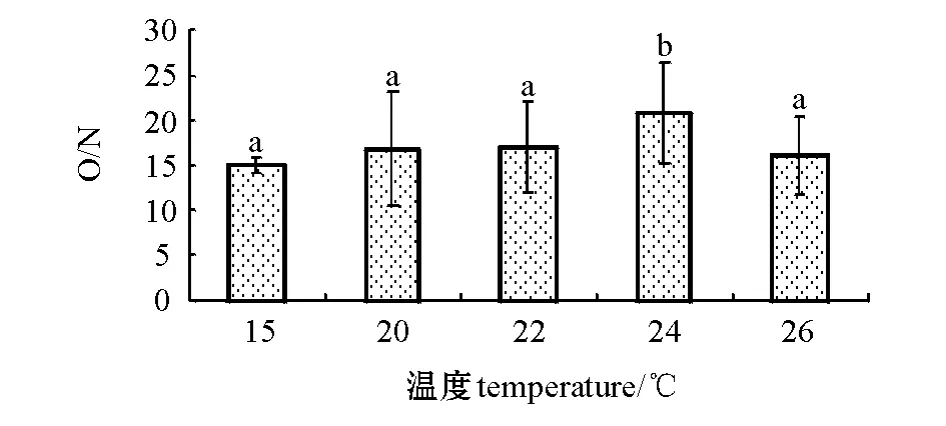
图4 不同温度下虾夷扇贝的氧氮比(O/N)Fig.4 The ratio of O to N in yesso scallop M.yessoensis at different temperature
从表1可见:15 ~20 ℃与20 ~22 ℃时,耗氧率和排氨率的Q10系数均最大,其中15 ~20 ℃时耗氧率和排氨率的Q10系数分别为3.476和2.770;20 ~22 ℃时,耗氧率和排氨率的Q10系数分别为5.079和8.920;而22 ~24 ℃与24 ~26 ℃时,耗氧率和排氨率的Q10系数均较小,仅为0.077 ~0.523。

表1 不同温度处理组虾夷扇贝耗氧率和排氨率的Q10 系数Tab.1 Q10 values for oxygen consumption rate and ammonia excretion rate in yesso scallop M.yessoensis at various temperature
3 讨论
3.1 不同温度下虾夷扇贝的存活率
虾夷扇贝是大型冷水性双壳贝类,早期文献记载虾夷扇贝的耐温上限为23 ℃,超过23 ℃时虾夷扇贝代谢活动缓慢,基本处于不摄食状态,从而导致机体消瘦,最后死亡,其中15 ℃是虾夷扇贝最适生长温度[4]。本研究发现,在缓慢升温试验过程中,24 ℃与26 ℃处理组虾夷扇贝的存活率最低,分别为56.31%和26.13%,其余处理组存活率均高于85%。在温度突变过程中,当温度从15℃突变到20 ~24 ℃时,经96 h 胁迫后,虾夷扇贝存活率均高于82.29%。但从15 ℃突变到26 ℃时则会引起虾夷扇贝大量死亡,参照生物毒性试验方法计算发现,96 h 虾夷扇贝的半致死温度为23.81℃,与早期文献[3-4]报道的23 ℃基本一致。但与徐东等[3]报道的结果略有差异。徐东等[3]对壳长为56.300 mm、湿质量为21.994 g、软体部干质量为0.935 g 的虾夷扇贝进行试验时发现,温度达到25 ℃时,虾夷扇贝未出现死亡现象,但摄食率非常低。本研究结果与徐东的研究结果存在差异的原因可能与试验所用虾夷扇贝个体大小有关,本试验中所用虾夷扇贝壳长为95.380 mm、湿体质量为107.580 g、软体部干质量为4.670 g,规格较大。另外,袁有宪等[5]通过试验也发现,个体小的栉孔扇贝Chlymys farreri 比个体大的耐高温程度要强一些。有关极限高温下,虾夷扇贝的个体大小对其存活率的影响还有待于进一步研究。
3.2 不同温度下虾夷扇贝的耗氧率和氨氮排泄率
目前,耗氧率和氨氮排泄率已经被认为是评估水产动物在不同胁迫环境中利用蛋白和能量的有效指标[6-9]。而温度可能是影响虾夷扇贝代谢率最明显的环境因子[10]。由简单能量平衡公式P=A-R(P 为生长能,A 为消化吸收能,R 为呼吸作用能)可知,如果假定饵料的消化吸收与温度无关,呼吸代谢升高则生长下降,呼吸代谢降低则生长增加[11]。本研究中发现,虾夷扇贝的耗氧率随温度的升高呈先升高后降低的趋势,15 ℃时虾夷扇贝的耗氧率显著低于20 ~24 ℃时,说明虾夷扇贝在15 ℃的环境中耗能少,更多的能量则用于生长,因此,15 ℃是虾夷扇贝最适生长温度,这与早期研究结果相一致[4]。26 ℃时,虽然虾夷扇贝耗氧率也较低,但已超过虾夷扇贝的耐温上限,其代谢活动非常微弱,基本处于不摄食状态,所以,虽然其耗氧率低,但生长依然缓慢。另外,虾夷扇贝的氨氮排泄率随温度升高表现为先增加后下降的趋势。这说明,在适宜温度下,温度升高能够加速蛋白和氨基酸的分解代谢,从而导致氨氮排泄升高。此外,本研究中虾夷扇贝耗氧率和氨氮排泄率随温度升高的变化趋势与双壳滤食性贝类中的墨西哥湾扇 贝Argopecten irradians concentricus[12]、栉 孔 扇贝[5]、硬壳蛤Mercenaria mercenaria[13]随温度的变化情况相似。
3.3 不同温度下虾夷扇贝的代谢水平
温度是影响贝类生理活动变化的重要因素之一,在适宜温度范围内,贝类代谢率随温度的升高而增高,超出此范围,代谢就会出现异常[14-16]。双壳贝类的Q10值一般为1.0 ~2.5,平均为2.0[17-19]。本研究结果显示,在试验温度范围内,虾夷扇贝耗氧率和排氨率的Q10值分别为0.077 ~3.467和0.123 ~8.920,超出一般结论值。这说明作为低温种的虾夷扇贝对温度变化较为敏感,适应温度变化的能力较弱。本研究结果与徐东等[3]的研究结果更为接近。
O/N 值是生物体内蛋白质与脂肪和碳水化合物分解代谢的比率,是动物代谢的一个重要指标。通过O/N 值能够估计动物代谢中能源物质的化学本质。Mayzalld[20]指出,如果机体完全由蛋白质提供能量,O/N 值约为7。Ikeda[21]认为,如果机体完全由蛋白质和脂肪氧化供能,O/N 值约为24。Conover等[22]表明,如果机体主要由脂肪或碳水化合物供能,O/N 值将会变为无穷大。本试验结果发现,虾夷扇贝的O/N 值为14.92 ~20.79,说明在试验设定的温度范围内,虾夷扇贝的代谢物质以蛋白质和脂肪氧化供能为主。试验结果与徐东等[3]报道的试验结果略有差异,笔者分析认为,造成差异的原因还是由于试验所用虾夷扇贝的大小不同所致。
[1]丁君,常亚青,张婧,等.虾夷扇贝“象牙白”品系与普通品系营养成分分析及评价[J].中国农业科技导报,2011,13(2):121-128.
[2]李文姬,薛真福.持续发展虾夷扇贝的健康增养殖[J].水产科学,2005,29(9):49-51.
[3]徐东,张继红,王文琪.温度变化对虾夷扇贝耗氧率和排氨率的影响[J].中国水产科学,2010,17(5):1101-1106.
[4]李文姬,谭克非.日本解决虾夷扇贝大规模死亡的启示[J].水产科学,2009,28(10):609-612.
[5]袁有宪,陈聚法,曲克明.栉孔扇贝对环境变化适应性研究——温度对存活、呼吸、摄食及消化的影响[J].中国水产科学,2000,7(3):24-27.
[6]Chen J C,Chia P G.Effects of unilateral eyestalk ablation on oxygen consumption and ammonia-N excretion of juvenile Penaeus japonicus Bate at different salinity levels[J].J Crustacean Biol,1995,15:434-443.
[7]Chen J C,Chen K W.Oxygen uptake and ammonia-N excretion of juvenile Penaeus japonicus during depuration following one-day exposure to different concentrations of saponin at different salinity levels[J].Aquaculture,1997,156:77-83.
[8]曹善茂,张丛尧,左骁.不同生态因子对曼氏皮海鞘耗氧率的影响[J].大连海洋大学学报,2012,27(3):210-214.
[9]刘青,宋莹莹,邢勇,等.温度和盐度对近亲真宽水蚤耗氧率与窒息点的影响[J].大连海洋大学学报,2012,27(3):205-209.
[10]Dall W,Hill B J,Rothlisberg P C.The Biology of the Penaeidae[M]//Blaxter J H S,Southward A J.Advances in Marine Biology.New York:Academic Press,1990.
[11]Dalla Via G J.Salinity responses of the juvenile penaeid shrimp Penaeus japonicus:I.Oxygen consumption and estimations of productivity[J].Aquaculture,1986,55:297-306.
[12]Mayzaud P.Respiration and nitrogen excretion of zooplankton:II Studies of the metabolic characteristics of starved animals[J].Mar Biol,1973,21:19-28.
[13]柴雪良,方军,林志华.温度对美国硬壳蛤滤食率、耗氧率和排氨率的影响[J].海洋科学,2005,29(8):33-36.
[14]常亚青,王子臣.皱纹盘鲍的个体能量收支[J].应用生态学报,1998,9(5):5l1-516.
[15]王俊,姜祖辉,唐启升.栉孔扇贝耗氧率和排氨率的研究[J].应用生态学报,2002,13(9):1157-1160.
[16]杨红生,张涛,王萍.温度对墨西哥湾扇贝耗氧率及排泄率的影响[J].海洋学报,1998,20(2):91-95.
[17]常亚青,王子臣.魁蚶耗氧率的初步研究[J].水产科学,1992,12:1-6.
[18]Clark B C.Ecological energetic of mussels Choromytilus meridionalis under simulated intertidal rock pool condition[J].J Exp Mar Biol Ecol,1990,137:63-77.
[19]Wilbur A E.Physiological energetics of the ribbed mussel Geukensia demissa(Dillwyn)in response to increased temperature[J].J Exp Mar Biol Ecol,1989,131:161-170.
[20]Mayzalld P.Respiration and nitrogen excretion of zooplankton:Ⅳ.The influence of starvation on the metabolism and biochemical composition of some species[J].Marine Biology,1976,37:47-58.
[21]Ikeda T.Nutrition ecology of marine zooplankton[J].Mem Fac Fish Hokkaido Univ,1974,22:1-77.
[22]Conover R J,Corner E D S.Respiration and nitrogen excretion by some marine zooplankton in relation to their life cycles[J].J Mar Biol Assoc U K,1968,48:49-75.
