黄颡鱼免疫球蛋白M基因的克隆与组织表达分析
2013-02-08叶仕根费阳春李强李华
叶仕根,费阳春,李强,李华
(大连海洋大学农业部北方海水增养殖重点实验室,辽宁大连 116023)
黄颡鱼免疫球蛋白M基因的克隆与组织表达分析
叶仕根,费阳春,李强,李华
(大连海洋大学农业部北方海水增养殖重点实验室,辽宁大连 116023)
根据GenBank中登录的免疫球蛋白M(IgM)基因编码氨基酸的保守序列以及鱼类密码子的偏好性设计简并引物,提取黄颡鱼Peltebagrus fulvidraco脾脏总RNA,经RT-PCR扩增首次获得了黄颡鱼IgM的部分序列 (675 bp)。以β-actin为内参基因,通过Real-time PCR法分析了黄颡鱼IgM基因的组织分布特点。结果表明,在黄颡鱼肝胰脏、脾脏、头肾、中肾、鳃、肌肉、皮肤和肠道中均检测到IgM基因的表达,头肾和脾脏是IgM表达的主要部位,肾脏、鳃、皮肤、肠道和肝胰脏等组织中表达量居中,肌肉中表达量最低。经鮰爱德华菌Edwardsiella ictaluri胞外产物免疫后,头肾、脾脏和肝胰脏中IgM基因表达呈现不同的变化规律。
黄颡鱼;IgM;克隆;组织分布
免疫球蛋白 (Immunoglobulins,Ig)是有颌类脊椎动物所特有的介导体液免疫的重要效应分子,在脊椎动物特异性免疫系统中起着关键作用。Ig分子包括重链和轻链两部分,根据重链恒定区化学结构的差异,可将Ig划分为多种类型。不同动物所拥有的Ig种类不同,目前已报道的硬骨鱼类Ig有IgM、IgD、IgZ/T和IgM-IgZ嵌合体等几种类型[1-5]。有报道指出,病原菌感染可明显提高鳜鱼IgM的表达水平,而IgD和IgZ无明显变化[6],内毒素刺激则可增加鲤IgM-IgZ嵌合体基因的转录[5],这说明不同类型的Ig有不同的免疫应答机制。在这些Ig分子中,IgM存在于所有有颌类脊椎动物中,其重链代表了从软骨鱼类到哺乳类的一个连续进化的过程[7]。同时,IgM也是鱼类最先表达的抗体,在体液免疫尤其是抵抗细菌性抗原侵扰方面起着重要作用[8-9]。目前,已有多种鱼类IgM重链基因被克隆鉴定,诸如模式鱼类斑马鱼Danio rerio[10]和一些重要的经济鱼类[11-17]。这些研究结果显示,不同鱼类的IgM重链之间存在基因数目差异和序列特异性,同时不同鱼类IgM基因的组织分布也存在差异。
黄颡鱼Peltebagrus fulvidraco是重要的经济鱼类之一,然而对其IgM基因的研究至今尚未见报道。本研究中,作者根据GenBank数据库中登录的黄颡鱼近缘物种的IgM重链序列信息,通过设计简并引物克隆IgM重链基因的部分片段,同时采用Real-time PCR方法研究黄颡鱼IgM重链基因在各组织中的表达情况以及经胞外产物 (OMPs)免疫刺激后的变化规律,以期为黄颡鱼的抗感染机制和免疫防治技术研究提供理论依据,对了解鱼类免疫应答的分子机制以及免疫系统的起源与进化等具有重要意义。
1 材料与方法
1.1 材料
试验用黄颡鱼体质量为35~50 g,购自辽宁营口某黄颡鱼养殖场,驯养一周后用于试验。
Trizol总RNA提取试剂盒、M-MLV反转录酶、荧光定量PCR试剂盒SYBRⓇGreen SuperMix-UDG试剂盒等购自上海 Invitrogen公司;pMD18-T载体、TaqDNA聚合酶、DNA Marker均购自宝生物(大连)工程有限公司;引物由北京六合华大基因科技股份有限公司合成。
1.2 方法
1.2.1 IgM重链基因核心序列的扩增 黄颡鱼暂养一周后,随机挑选2尾,取其脾脏,按照试剂盒说明书方法提取黄颡鱼脾脏总RNA。选用经电泳和紫外分析检测条带清晰、完整性好的RNA用于后续的反转录和PCR扩增。取4 μL(约2 μg)总RNA悬液,使用M-MLV反转录酶和oligo(dT)18于37℃下进行反转录,所得cDNA于-20℃下保存备用。根据NCBI中已登录的鱼类免疫球蛋白重链恒定 (CH)区氨基酸保守序列设计简并引物[13],正向引物 F'为 5'CCNACNCARACNGAYATHGAY 3',反向引物R'为5'NYTNACNTGYTAYGTNAARGA 3',其中N为G、A、T或C;Y为T或C;R为G或A,W为A或T。在此基础上,参照鱼类密码子偏好性[18]对引物做进一步修改,去掉一些鱼类较少使用的密码子,针对性地降低引物的简并度。修改后的正向引物IgM-F为5'CCAACCCAAACAGAMATAGAC 3',反向引物IgM-R为5'TCTTTWACATAGCAAGTCAGG 3',引物简并度由1 536、4 096下降至2。以cDNA第一链为模板进行PCR扩增,将所得目的条带回收纯化并与PMD-18T载体连接,转化至DH5α中,经菌落PCR和酶切鉴定后,送北京六合华大基因科技股份有限公司测序。
1.2.2 序列分析 参照李颖等[19]的方法,将测序得到的序列在NCBI网站(http://www.ncbi.nlm.nih.gov/blast)进行相似性搜索和比对,并提交GenBank。采用Expasy网站的Prosite在线工具(http://prosite.expasy.org/prosite.html)进行编码蛋白质的保守结构域分析,用Clustal X1.8和DNA 6.0程序进行氨基酸序列同源性分析和作图。
1.2.3 IgM重链基因组织表达量的检测 随机选取3尾黄颡鱼,分别取肝胰脏、脾脏、头肾、肾脏、鳃、肌肉、皮肤和肠等8个组织,液氮速冻后按上述方法提取各组织总RNA并反转录得到cDNA。根据“1.2.1”和“1.2.2”节中获得的IgM重链基因序列设计实时定量PCR引物,正向引物 IgM-F1为 5'AGAGCCAGAAGTGAGCATTA 3',反向引物IgM-R1为5'CTTGGCAGGTGTATGTGG 3'。根据黄颡鱼β-actin基因的序列 (GenBank登录号:EU161066.1)设计实时定量PCR内参引物,正向引物 ACTIN-F为5'GATCCGGTATGTGCAAGGCT 3',反向引物 ACTIN-R为 5'TGCCAGATCTTCTCCATATCA 3'(表1)。引物合成后,以cDNA第一链为模板进行PCR扩增,经E.B染色,用15 g/L琼脂糖电泳检验其特异性。

表1 基因克隆与组织表达分析所用引物序列Tab.1 Sequences of the primers used in the gene cloning and tissue expression
荧光定量PCR参照刘俊等[20]的方法,并作适当修改。采用2—△△CT法计算基因在各组织中的表达量:
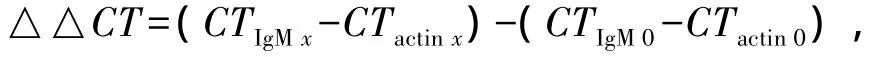
其中:CT为样品管中荧光强度达到特定阈值时的扩增循环数;x表示8个待测组织中的任一组织;0表示8个待测组织中相对表达量最低的组织。用SYBR Green qPCR试剂盒,在Step one定量PCR仪上进行实时定量扩增。首先将反转录获得的cDNA模板以及引物浓度进行优化,将CT值调整为20左右。扩增反应程序为:95℃下预变性2 min;95℃下变性30 s,62℃下退火30 s,共进行40个循环。每个样品均进行IgM和β-actin表达量的测定。每个样本同时设立3个平行管,每次反应均设置熔解曲线分析,以验证扩增反应的特异性。反应完成后,系统软件将自动给出每个样本扩增的IgM和β-actin基因的CT值,在此基础上进一步进行基因表达差异的分析。
1.2.4 鮰爱德华菌OMPs免疫对IgM基因表达的影响 取大连海洋大学农业部北方海水增养殖重点实验室分离保存的黄颡鱼病原菌鮰爱德华菌A86,经扩大培养后,参照李强等[21]的方法提取OMPs。调整OMPs浓度为1 mg/mL,与弗氏完全佐剂等体积混合后腹腔注射免疫黄颡鱼30尾 (100 μL/尾)。分别于免疫接种后第4、8、14、21和28天随机选取3尾鱼,取其肝胰脏、脾脏和头肾组织,液氮速冻后按照上述方法提取各组织总RNA并反转录得到cDNA。另随机选取3尾未免疫鱼,取其肝脏、脾脏和头肾组织作为免疫前对照。采用IgM-F1/R1进行IgM基因表达的实时定量扩增,ACTIN-F/R用作内参基因扩增引物,试验方法和数据计算同“1.2.3”节。
2 结果
2.1 脾脏RNA质量和定量PCR引物特异性验证
黄颡鱼脾脏总RNA经甲醛变性凝胶电泳分析,可见28S RNA和18S RNA两条清晰条带,亮度在2∶1左右,用核酸紫外分析仪检测样品的OD260nm/OD280nm值为1.8~2.0,表明RNA质量较好,无蛋白质、酚等污染(图1)。对黄颡鱼IgM基因表达分析引物IgM-F1/R1和内参基因β-actin引物ACTIN-F/R扩增产物的熔解曲线分析表明,均呈现单一峰值 (84.38、84.97℃),说明PCR产物单一,无非特异性扩增,引物特异性强(图2)。

图1 黄颡鱼脾脏总RNA琼脂糖凝胶电泳Fig.1 Agarose gel electrophoresis of the total RNA in spleen of the yellow catfish
2.2 黄颡鱼IgM重链基因恒定区的基因克隆
以脾脏总RNA反转录得到的cDNA为模板,以IgM-F/R为引物进行PCR扩增,扩增产物经凝胶电泳检测得到一条约为700 bp且与预期扩增片段大小相符的条带 (图3)。条带经切胶回收后连接pMD-18T载体,再经PCR扩增和酶切鉴定初步确认后送交测序,最终获得一段675 bp的序列并提交GenBank(GenBank登录号:JQ067604.1)。
2.3 黄颡鱼IgM重链基因恒定区的序列分析
分析发现,黄颡鱼IgM重链基因恒定区序列共编码225个氨基酸残基 (GenBank登录号:AEY79771),包含4个保守的半胱氨酸残基 (图4,第15、74、120、172位,加框表示)。进一步分析发现,黄颡鱼IgM重链基因恒定区第15、74、120、172位半胱氨酸残基的分布符合IgC(免疫球蛋白恒定区)结构域的特征(C-X(n)-C,X为任意氨基酸残基),可分别形成两个二硫键,组成两个包含89个氨基酸残基的 IgC结构域 (图4,第1~89位和第98~176位氨基酸残基,下划线表示)。经Blast搜索和序列比对分析显示,其编码氨基酸与瓦式黄颡鱼、大鳍鳠、斑点叉尾鮰、草鱼和斑马鱼等鱼类IgM重链基因编码氨基酸序列的相似度为56% ~88%(表2,图5),与鱼类IgM基因具有较高同源性。

表2 黄颡鱼IgM部分序列与其他鱼类IgM氨基酸序列的一致性和相似度比较Tab.2 Similarity and identity of deduced amino acid sequence of IgM in yellow catfish Peltebagrus fulvidraco,with other fishes
2.4 黄颡鱼IgM基因的组织表达
以β-actin为内参基因,黄颡鱼肝胰脏、脾脏、头肾、肾脏、鳃、肌肉、皮肤和肠道等8个组织中IgM基因表达情况如图6所示。从图6可见,IgM基因在黄颡鱼各检测组织中均有表达,但各组织间的表达水平存在较大差异。各组织中表达量由高到低依次为头肾、脾脏、肾脏、鳃、皮肤、肠道、肝胰脏和肌肉,头肾和脾脏中IgM基因的表达量明显高于其他组织,分别为表达量最低的肌肉组织的10.3和10.1倍;肾脏、鳃、皮肤、肠和肝胰脏中IgM基因的表达量居中,分别为肌肉组织的5.3、3.7、3.0、2.7和2.2倍。
2.5 OMPs免疫对黄颡鱼IgM基因表达的影响
由于样品保存不当 (从液氮取出时,装有样品的EP管因温差过大炸裂),免疫前的黄颡鱼肝胰脏、脾脏和头肾组织样品未能用于IgM基因表达的定量检测。因此,各组织IgM基因表达变化情况均采用免疫后 (第4天)首次取样为初始表达水平,并以此为基准进行比较。以β-actin为内参基因,经OMPs免疫后,黄颡鱼肝胰脏、脾脏和头肾组织 (使用专用冻存管保存于液氮中)IgM基因的表达情况如图7所示。从图7可见:黄颡鱼肝胰脏、脾脏和头肾中IgM基因的表达呈现出不同的变化规律,与脾脏和头肾相比,肝胰脏中IgM基因表达在整个取样过程中变化不太明显,最高表达(免疫后第21天)和最低表达 (第14天)分别为初始水平的1.06、0.81倍;3个组织中,头肾和脾脏中IgM基因表达变化规律较为相似,均为短暂2.90和1.92~2.21倍。下降 (第8天)后再升高,最高表达分别出现在第14天和21天;脾脏IgM基因表达的最高值和最低值分别为初始水平的2.90和0.71倍,头肾IgM基因表达的最高值和最低值分别为初始水平的2.21和0.82倍;脾脏和头肾组织中IgM基因表达自免疫后第14天到第28天试验结束时均维持了较高的表达水平,分别为初始表达水平的1.84~

图2 Real-time PCR扩增产物的熔解曲线分析Fig.2 Melting curves of the PCR products of IgM-F1/R1 and ACTIN-F/R

图3 用简并引物扩增黄颡鱼IgM序列的PCR产物电泳Fig.3 Agarose gel electrophoresis of PCR products of IgM sequence by degenerate primers in yellow catfish

图4 黄颡鱼IgM部分序列编码氨基酸的分析Fig.4 Deduced amino acid sequences by IgM from yellow catfish Peltebagrus fulvidraco

图5 黄颡鱼IgM部分序列与其他鱼类IgM重链氨基酸序列的比较分析Fig.5 Comparison of multiple amino acid sequences of IgM in yellow catfish Peltebagrus fulvidraco,with other fishes

图6 Real-time PCR检测黄颡鱼IgM基因的组织分布Fig.6 Expression levels of IgM gene in various tissues detected by Real-time PCR

图7 OMPs免疫对黄颡鱼各组织IgM表达的影响Fig.7 Expression levels of IgM gene in various tissues of yellow catfish challenged by OMPs detected by Real-time PCR
3 讨论
本研究中,通过RT-PCR方法首次成功克隆了黄颡鱼IgM重链基因恒定区部分cDNA序列。该序列编码氨基酸中包含4个保守的半胱氨酸残基,可分别形成两个二硫键,组成两个IgC(Ig恒定区)结构域。同源性分析发现,其编码氨基酸与瓦式黄颡鱼、大鳍鳠、斑点叉尾鮰、草鱼和斑马鱼等鱼类的IgM重链基因编码氨基酸序列的相似度为56%~88%,具有较高的同源性。在获得黄颡鱼IgM重链基因部分序列的基础上,采用Real-time PCR方法检测了IgM重链基因在黄颡鱼不同组织中的表达情况以及经OMPs免疫后其在各组织中表达的变化规律,为黄颡鱼乃至其他鱼类的抗感染机制和疾病防治研究等提供了理论依据。
在新基因的克隆中,根据近缘物种相关蛋白氨基酸序列的保守区域设计简并引物是常用的方法之一。但由于密码子的简并性和摇摆性,常常使得设计出来的引物简并度非常高,不利于PCR扩增反应的进行。张晓峰等[18]研究发现,鱼类密码子与其他生物一样具有明显的偏好性,即编码同一种氨基酸的密码子在翻译成蛋白质时并非均一使用,通常是某些密码子被优先使用,某一物种或某一基因通常会倾向于使用一种或几种特定的同义密码子,即所谓的最优密码子,此现象被称为密码子偏性。本研究中,参照鱼类密码子偏好性对引物进行了针对性修改,使两条引物的简并度分别由1 536、4 096下降至2,提高了引物的扩增效率,成功克隆到黄颡鱼IgM基因重链恒定区的部分cDNA序列。
Real-time PCR分析表明,黄颡鱼头肾和脾脏是IgM基因mRNA的主要分布组织,其表达量显著高于其他组织。本研究结果与李春涛等[22]对大鳍鳠以及王欣欣等[23]对草鱼IgM基因mRNA分布情况的研究结果类似。胡瑜兰[24]和彭博[25]研究发现,斑马鱼和鲫的头肾中免疫球蛋白具有较高表达水平。头肾和脾脏中较高的IgM基因mRNA或免疫球蛋白表达水平与其抗体产生细胞发生的主要器官的功能是相适应的,这从侧面证实了头肾和脾脏是鱼类免疫的主要场所[26-27]。值得注意是,作为鱼类造血器官之一,黄颡鱼肾脏中也有较高的IgM基因mRNA表达水平,为头肾和脾脏的50%左右。但这与欧洲鳗鲡[16]和大鳍鳠[22]肾脏中IgM基因mRNA表达水平高于脾脏中的研究结果有一定差异。这可能与物种差异、动物机体状况、环境等因素有关,如季节变化会影响鱼体内免疫球蛋白的表达水平,夏季高于冬季;运输会影响斑点叉尾鮰对抗原刺激的应答反应[28-29]。上述结果表明,头肾、脾脏和肾脏是鱼类IgM基因mRNA和免疫球蛋白表达的主要场所。此外,本研究中还发现,黄颡鱼肌肉组织中IgM基因mRNA的表达水平是所有被检测组织中最低的,这与对欧洲鳗鲡的研究结果一致。肌肉组织中IgM低表达可能与肌肉组织中MHC表达量较低有关[30]。因为MHC在免疫应答的启动和免疫调节中发挥重要作用,肌肉组织中较低的MHC表达量使得其自身难以具备产生免疫反应的分子基础[31]。
由于样本保存的原因,本研究中免疫前样品未能用于IgM基因表达的定量分析。研究表明,鳜和大鳍鳠经嗜水气单胞菌灭活疫苗免疫后以及剑尾鱼经溶藻弧菌灭活疫苗免疫后,其IgM基因表达水平在第6~10天后显著升高,但在前期如第3天 (剑尾鱼)、第4天 (鳜)和第5天 (大鳍鳠)变化并不十分明显[22,32-33]。因此,本研究中免疫后 (第4、8、14、21天)样品中的IgM基因表达情况仍能反映出其变化规律。经OMPs免疫后,黄颡鱼脾脏和头肾中IgM基因表达变化趋势相似,均为先小幅降低 (第8天)后升高,且自第14天起维持在较高表达水平。这与嗜水气单胞菌灭活疫苗免疫后,大鳍鳠脾脏和头肾中IgM基因表达持续升高并维持在较高的表达水平的结果一致[22]。但与鳜经嗜水气单胞菌灭活疫苗免疫后,头肾和脾脏中转录水平峰值出现在第7天,至第28天时鳜头肾中转录水平下降到免疫前水平而脾脏仍维持在较高表达水平的研究结果有一定差异[33]。这可能与物种和取样时间长短有一定关系。Zilberg等[34]研究发现,对斑点叉尾鮰注射鮰爱德华菌后,其血细胞、脾脏和头肾中IgM基因表达量在13 d内均显著增加。Raida等[35]研究发现,对虹鳟注射鲁氏耶尔森氏菌Yersinia ruckeri后,其脾脏和头肾中IgM基因表达量在21 d内都有增加。这些结果表明,鱼类IgM分子在3周内可以识别细菌抗原[6]。尽管在未经免疫刺激的情况下,肝胰脏中IgM基因表达水平较低 (仅为头肾或脾脏的1/4左右),作为鱼类重要的代谢器官,肝脏组成细胞参与肝脏免疫调节[36],其仍然可能在免疫防御中发挥重要作用。然而,本研究结果表明,经OMPs免疫后黄颡鱼肝胰脏中IgM基因表达量无明显变化。作者的另一研究发现,经鮰爱德华菌OMPs免疫可显著提升黄颡鱼肝胰脏中CAT、AKP和SOD活性 (另文发表)。这些结果表明,鱼类特异性免疫防御更多的是与脾脏、头肾等组织相关,肝胰脏更多的是通过增加一些免疫酶的活性来发挥免疫防御作用,主要参与机体的非特异性免疫防御,但其详细机制仍有待于进一步探讨。
综上所述,本研究中成功克隆到了黄颡鱼IgM基因的部分序列,并检测了其组织分布模式和经OMPs免疫后的变化规律,研究结果将有助于对黄颡鱼IgM结构与功能的理解,为鱼类的免疫防治技术和分子免疫应答机制研究提供了基础资料。
[1] Pilstrom L,Bengten E.Immunoglobulin in fish-genes,expression and structure[J].Fish & Shellfish Immunol,1996,6:243-262.
[2] Saha N R,Suetake H,Kikuchi K,et al.Fugu immunoglobulin D:a highly unusual gene with unprecedented duplications in its constant region[J].Immunogenetics,2004,56:438-447.
[3] Danilova N,Bussmann J,Jekosch K,et al.The immunoglobulin heavy-chain locus in zebrafish:identification and expression of a previously unknown isotype,immunoglobulin Z[J].Nat Immnol,2005,6(3):295-302.
[4] Hansen J D,Landis E D,Phillips R B.Discovery of a unique Ig heavy-chain isotype(IgT)in rainbow trout:implications for a distinctive B cell developmental pathway in teleost fish[J].Proc Natl Acad Sci USA,2005,102:6919-6924.
[5] Savan R,Aman A,Nakao M,et al.Discovery of a novel immunoglobulin heavy chain gene chimera from common carp(Cyprinus carpio L.)[J].Immunogenetics,2005,57:458-463.
[6] Tian J Y,Sun B J,Luo Y P,et al.Distribution of IgM,IgD and IgZ in mandarin fish,Siniperca chuatsi lymphoid tissues and their transcriptional changes after Flavobacterium columnare stimulation[J].Aquaculture,2009,288:14-21.
[7] Wilson M R,Warr G W.Fish immunoglobulins and the genes that encode them[J].Ann Rev Fish Dis,1992,2:201-221.
[8] Reddy P S,Corley R B.The contribution of ER quality control to the biologic functions of secretory IgM[J].Immunol Today,1999,20:582-588.
[9] Klimovich V B,Samoilovich M P,Klimovich B V.Problem of J-chain of immunoglobulins[J].Journal of Evolutionary Biochemistry and Physiology,2008,44(2):151-166.
[10] Danilova N,Hohman V S,Kim E H,et al.Immunoglobulin variable-region diversity in the zebra fish[J].Immunogenetics,2000,52:81-91.
[11] Ghaffari S H,Lobb C J.Cloning and sequence analysis of channel catfish heavy chain cDNA indicate phylogenetic diversity within the IgM immunoglobulin family[J].J Immunol,1989,142:1356-1365.
[12] Hansen J,Leong J A,Kaattari S.Complete nucleotide sequence of a rainbow trout cDNA encoding a membrane-bound form of immunoglobulin heavy chain[J].Mol Immnol,1994,31(6):499 -501.
[13] Nakao M,Moritomo T,Tomana M.Isolation of cDNA encoding the constant region of the immunoglobulin heavy-chain from common carp(Cyprinus carpio L.)[J].Fish & Shellfish Immunol,1998,8:425-434.
[14] Zhang Y A,Nie P,Wang Y P.cDNA sequence encoding immunoglobulin M heavy chain of the mandarin fish Siniperca chuatsi[J].Fish & Shellfish Immunol,2003,14:477-480.
[15] Cheng C A,John J A C,Wu M S,et al.Characterization of serum immunoglobulin M of grouper and cDNA cloning of its heavy chain[J].Veterinary Immunology and Immunopathology,2006,109:255-265.
[16] Feng J J,Guan R Z,Lin P,et al.Molecular cloning and characterization analysis of immumoglobulin M heavy chain gene in European eel(Anguilla anguilla)[J].Veterinary Immunology and Immunopathology,2009,127:144 -147.
[17] 冯汉如.南方鲇IgM重链基因cDNA克隆及该基因在小瓜虫感染下的表达特征分析[D].重庆:西南大学,2010.
[18] 张晓峰,孙效文.鲤鱼和斑马鱼同义密码子使用偏性分析[J].水产学杂志,2010,23(4):23-29.
[19] 李颖,周一兵,万良,等.双齿围沙蚕Hsp70 cDNA基因的克隆及序列分析[J].大连海洋大学学报,2012,27(6):502-507.
[20] 刘俊,赵金良,张敏,等.鳜胰岛素样生长因子-Ⅱ cDNA基因的克隆与表达特征[J].大连海洋大学学报,2012,27(6):11-17.
[21] 李强,刘海燕,黄华,等.鮰爱德华菌黄颡鱼分离株外膜蛋白的抗原性分析[J].广东海洋大学学报,2011,31(3):85-89.
[22] 李春涛,张其中,杨莹莹,等.大鳍鳠免疫球蛋白 M重链基因的克隆及表达分析[J].水产学报,2011,35(11):1684-1693.
[23] 王欣欣,孙宝剑,昌鸣先,等.草鱼免疫球蛋白M重链基因的克隆及表达[J].水产学报,2008,32(1):13-20.
[24] 胡瑜兰.硬骨鱼类补体关键因子C1q及免疫球蛋白分子克隆、进化和功能的初步研究[D].杭州:浙江大学,2009.
[25] 彭博.鲫鱼免疫球蛋白基因的鉴定、应答和功能研究[D].杭州:浙江大学,2008.
[26] 孟庆闻,苏锦祥,李婉端.鱼类比较解剖[M].北京:科学出版社,1987:361-363.
[27] 马燕梅,林树根,王全溪,等.花鲈头肾的显微结构和超显微结构[J].福建农林大学学报:自然科学版,2008,37(2):190-193.
[28] Ellsaesser C F,Clem L W.Hematological and immunological changes in channel catfish stressed by handling and transport[J].J Fish Biol,1986,28:511-521.
[29] Nakanishi T.Seasonal changes in the immune responses and lymphoid tissue on the marine teleost,Sebastiscus marmoratus[J].Vet Immunol Immunopathol,1986,12:213-222.
[30] Koppang E O,Hordvik I,Bjerekas I,et al.Production of rabbit antisera against recombinant MHC classⅡ:chain and identification of immunoreactive cells in Atlantic salmon(Salmo salar)[J].Fish & Shellfish Immunol,2003,14(2):115-132.
[31] 冯建军,关瑞章,林鹏,等.欧洲鳗鲡免疫球蛋白M重链基因的原核表达与不同组织中表达变化的定量分析[J].华中农业大学学报,2009,28(6):719-725.
[32] 韩进刚,付小哲,石存斌,等.剑尾鱼IgM基因的克隆及免疫对其组织表达的影晌[J].广东海洋大学学报,2007,27(6):1-6.
[33] 刘雨果,潘厚军,陈偿,等.嗜水气单胞菌灭活疫苗浸泡后鳜IgM基因表达量和抗体效价的变化[J].淡水渔业,2009,36(6):41-46.
[34] Zilberg D,Klesius P H.Quantification of immunoglobulin in the serum and mucus of channel catfish at different ages and following infection with Edwardsiella ictaluri[J].Veterinary Immunology and Immunopathology,1997,58:171-180.
[35] Raida M K,Buchmann K.Temperature-dependent expression of immune-relevant genes in rainbow trout following Yersinia ruckeri vaccination[J].Diseases of Aquatic Organisms,2007,77:41 -52.
[36] 张云霞,刘杞.肝窦内皮细胞与免疫耐受[J].世界华人消化杂志,2006,14(28):2776-2779.
Molecular cloning and expression of partial IgM cDNA sequence in different tissues of yellow catfish Peltebagrus fulvidraco
YE Shi-gen,FEI Yang-chun,LI Qiang,LI Hua
(Key Laboratory of Mariculture& Stock Enhancement in North China's Sea,Ministry of Agriculture,Dalian Ocean University,Dalian 116023,China)
Partial cDNA sequence(675 base pairs)was cloned in total DNA of spleen in yellow catfishPeltebagrus fulvidracoby degenerated primers designed based on fish IgM heavy chain gene in GenBank and by RT-PCR.The expression of IgM gene in different tissues of the yellow catfish was studied by real time PCR with internal control β-actin.The results showed that the IgM gene was constitutively expressed in all detected tissues including hepatopancreas,spleen,head kidney,kidney,gill,skin,intestine and muscle,the maximal expression levels in spleen and head kidney,the minimal expression level in muscle,and the mid-level in the other tissues.There was a different expression pattern of the IgM gene in head kidney,spleen,and hepatopancreas in the yellow catfish challenged with ecto-products of Edwardsiella ictaluri.
Peltebagrus fulvidraco;immunoglobulin M;clone;tissue distribution
Q786
A
2013-03-13
国家“十二五”科技支撑计划项目 (2011BAD13B03)
叶仕根 (1976-),男,副教授。E-mail:yedlsy@dlou.edu.cn通信作者:李华 (1958-),女,教授。E-mail:lihua@dlou.edu.cn
2095-1388(2013)06-0515-07
