不同特质焦虑水平的选择性注意偏向*
2013-01-31彭家欣杨奇伟罗跃嘉
彭家欣 杨奇伟 罗跃嘉
(1北京师范大学,认知神经科学与学习国家重点实验室,北京 100875)(2深圳大学 情绪与社会认知科学研究所,深圳 518060)
1 前言
焦虑是对未来威胁和不幸的忧虑预期,并伴随着紧张的烦躁不安或一定的身体症状(Lang,Davis,&Öhman,2000)。适当程度的焦虑水平能够提高个体的警惕性,使其更快地知觉和处理环境中的潜在威胁,因此具有一定的生存适应意义。然而如果个体的焦虑状态长期处于较高水平,则可能会演变为焦虑障碍。焦虑障碍是最常见、最普遍的精神障碍,约 20%~30%的正常人在一生中的某段时间会受到焦虑障碍的困扰(Kessler et al.,1994)。因此,与焦虑相关的研究在心理健康研究领域中一直得到广泛的关注。
高特质焦虑的人群在一般情况下(没有特别的焦虑诱发事件)也保持着较高的焦虑水平,是临床焦虑障碍的“易感人群”(McNally,2002)。先前的行为研究的结果一致表明,焦虑患者常常对环境中的负性情绪信息表现出选择性注意的倾向。例如研究者采用“点探测”任务发现广泛性焦虑障碍、社会焦虑和特质焦虑的个体对代替威胁性面孔的目标的反应要快于代替中性面孔的目标,说明了焦虑障碍者对威胁信息更加警惕(Bradley,Mogg,White,Groom,&Bono,1999;Mogg &Bradley,2002);一项采用“视觉搜索”任务检测焦虑患者注意障碍特点的研究发现,社会焦虑个体相对于非焦虑个体在中性面孔的背景下,对愤怒面孔的反应时快于对愉快面孔的反应时,说明社会焦虑个体能够更快觉察到威胁性信息(Gilboa-Schechtman,Foa,&Amir,1999);国内学者也采用“视觉搜索”任务发现,高特质焦虑者不管是在平静的状态下,还是焦虑的状态下,在恐惧面孔的背景下对中性面孔的反应显著慢于快乐面孔的背景下的反应,说明了高特质焦虑者难以将注意从威胁性刺激中脱离出来(高鹏程,黄敏儿,2008)。这些研究均证实了对威胁性刺激的选择性注意偏向在不同的焦虑类型、不同的实验范式下广泛存在,表现为高焦虑者对威胁信息更加警惕、反应更加敏捷并且难以脱离,说明了高焦虑者对威胁性刺激投入了更多的注意资源通过对150多篇关于焦虑研究进行元分析后发现,高特质焦虑者和临床焦虑者均表现出对威胁信息的选择性注意偏向,并且这种偏向的大小在两者间没有显著的差别,研究者由此认为对威胁信息的选择性注意偏向是高特质焦虑者容易发展成临床焦虑的重要原因。因此,本研究通过比较高低特质焦虑的个体,试图揭示高焦虑者对威胁性刺激的选择性注意偏向的神经机制(Bar-Haim,Lamy,Pergamin,Bakermans-Kranenburg,&van IJzendoorn,2007)。
大量的行为研究均证实了高焦虑个体存在对威胁性信息注意偏向的特点,然而这种选择性注意偏向发生在什么时候,其神经反应是怎么样的?目前仍然没有得到一致的结论。由于 ERP(event related potential)能够直接地并且精确到毫秒级地实时得记录神经元活动(Hajcak,MacNamara,&Olvet,2010),具有较高的时间分辨率,因此能够提供行为学实验难以提供的精确时程方面和神经反应的信息。近年来越来越多的研究者使用 ERP技术来研究焦虑个体在情绪加工时注意分配的特点(Bar-Haim,Lamy,&Glickman,2005;Li,X.Y.,Li,X.B.,&Luo,2005;Li,Wang,Poliakoff,&Luo,2007;MacNamara &Hajcak,2009)。例如Bar-Haim等人(2005)的研究发现高特质焦虑的个体在完成注意转移任务时,相对于线索为中性面孔时,当线索为愤怒面孔(与威胁有关)时诱发了一个更加大的P2,而低特质焦虑个体并没有这种注意偏向,说明了高特质焦虑的个体对威胁性面孔投入了比较多的注意(Bar-Haim et al.,2005)。然而 Holmes,Nielsen和Green(2008)却在高、低特质焦虑的个体中均发现了这种 P2效应,即高、低特质焦虑的个体同样对威胁有关的面孔投入了更多的注意资源(Holmes et al.,2008)。同样的矛盾情况也在早期的知觉成分P1上出现,例如 Holmes等(2008)发现高特质焦虑的个体观看恐惧面孔时比中性面孔诱发了更大的 P1,尽管这种注意偏好的效应只在左侧上发现;然而,Li等人(2005)发现高低焦虑的个体都出现威胁图片诱发更大P1的现象。
行为研究的结果一致发现了高特质焦虑者对威胁性刺激的选择性注意偏向,但是 ERP的研究却发现了相互矛盾的结果。这种不一致的其中一个重要原因很可能是ERP的成分P1、P2很容易受到不同刺激知觉特征差异的影响(Hajcak,Weinberg,MacNamara,&Foti,2012)。例如以P1成分为例,由于不同情绪面孔之间的知觉差异较小,由于高特质焦虑者的选择性注意偏向,因此只在高焦虑组中发现 P1效应,而威胁的情绪图片(常常是危险动物)和中性图片(常常是普通物品)情景轮廓等特征差异比较大,因此高低特质焦虑的个体都很容易辨别这两种刺激,都出现P1效应(Li et al.,2005)。那么,在复杂的情境中,高特质焦虑者对威胁性刺激的选择性注意偏向究竟发生在什么时候? 已有的研究发现,在P1之后的ERP另一成分N1,对视觉刺激的情绪内容敏感,负性图片比起中性图片诱发了一个更大的N1(Foti,Hajcak,&Dien,2009;Weinberg &Hajcak,2010),增大的 N1说明了个体对刺激进行知觉加工时投入了更多的注意资源(Mangun &Hillyard,1990)。因此,N1可能是高特质焦虑者进行情绪图片加工时出现的对威胁图片选择性注意偏向的可靠指标。
综上所述,本研究试图采用 ERP技术,以 N1成分为主要指标来考察高低特质焦虑的个体在加工复杂情景下的情绪图片时的选择性注意偏向的神经反应的差异。通过上述的文献回顾,我们假设:1)高特质焦虑者对恐惧图片刺激比较敏感,分配更多的注意资源,相对于中性图片诱发出更负的 N1;2) 低特质焦虑者观看恐惧图片和中性图片诱发的N1并没有显著的差异。尽管 N2、P3和慢波(SW)并非本研究关注的重点,但是已有的研究发现,这些成分与情绪加工密切相关(Dennis &Chen,2009;Foti &Hajcak,2008;Luo,W.,Feng,He,Wang,&Luo,Y.,-J.,2010),因此本研究也将这次成分纳入后续的分析中,以便更加全面地考察不同特质焦虑水平加工情绪图片时的选择性注意偏向。
2 方法
2.1 被试
采用状态-特质焦虑问卷(Spielbergerg Trait Anxiety Inventory,STAI)(Shek,1993;Spielberger,Gorsuch,Lushene,Vagg,&Jacobs,1983)中的“特质焦虑问卷”(STAI-T)测查了70名在校学生,将被试的问卷得分按照由高到低的顺序排列,取得分在前25%的被试为高特质焦虑组,得分在后 25%的被试为低特质焦虑组,最后得到高特质焦虑和低特质焦虑的个体各17人,其中低焦虑组女7人,男10人,高焦虑组女11人,男6人。34名被试平均年龄为21.94 ± 2.28岁。低特质焦虑组平均分为 35.29 ±4.22,高特质焦虑组为47.06 ± 3.91,t检验结果表明两组被试特质焦虑得分差异显著(t
=−8.42,p
<0.01)。被试的视力或者矫正后的视力正常。2.2 实验材料
从中国情绪图片系统(CAPS,白露,马慧,黄宇霞,罗跃嘉,2005)中选取效价在2~3分之间(1~9分,分数越低表示越消极,5分代表中性,分数越靠近9代表越积极)的恐惧动物图片31张和效价在5左右的物品图片 79张,另从国际情绪图片系统(IAPS,Lang,Bradley,&Cuthbert,1999)挑选取效价在 2~3分之间恐惧动物的图片 48张,通过photoshop CS 5处理成相同的大小和亮度后,另请21名在校学生分别对这 158张图片对其唤醒度和效价进行评定,最后得到 30张恐惧动物图片(效价2.63±0.47,唤醒度为7.29±0.46)和30张中性物品图片(效价为 5.02±0.17,唤醒度为 3.78±0.37)。t检验的结果表明恐惧图片和中性图片在效价(t
=25.67,p
<0.01)和唤醒度(t
=232.56,p
<0.01)的差异均显著。2.3 实验任务和程序
本研究使用 E-prime 2.0来控制和呈现所用刺激。本研究采用的实验范式改编自Holmes等(2008)的研究,被试的任务是:当前图片与上一张是同一张时,要求尽快做按键反应。与 Holmes等(2008)不同的是,本研究把图片刺激由原来的混合呈现改成 block呈现。本实验为系列研究的第一个,后续研究还需要对每类情绪图片的评定,采用 block呈现刺激的方式只需要每个 block后评价一次,可以节省时间,以及为了让实验间可以相互比较。本实验将30张恐惧图片和30张中性图片各呈现2遍,并且每类图片分成3个block,每个block 20张图片,共形成6个block共120个trials。每个trial的先后顺序按照伪随机的方法处理,以保证除了每个block中有 3~4张图片被连续呈现两次,其余图片两次呈现的间隔至少为4个trials。同一图片连续出现时,被试只在第二次出现时需要进行按键反应(共20个trials,不分析),其他情况只需要被动观看(100 trials)。
被试进入灯光控制的实验室后,舒服地坐在距离电脑屏幕75 cm的扶椅,并带上EEG电极帽,然后给被试一个详细的任务说明。在正式实验前,被试需要先进行练习,直到完全理解后才进入正式实验。正式实验的流程如图1,首先在白色屏幕中呈现一个黑色的“+”注视点(500 ms),提示一个trial即将开始,然后呈现 300~500 ms白色屏幕后出现一张图片(2000 ms),图片消失后出现1000 ms的白色屏幕,至此第一个 trial结束,再次出现黑色的“+”注视点(500 ms)时提示被试第二个 trial已经开始,被试需要从第二个 trial开始呈现的图片与第一个trial呈现的图片做比较,如果是同一张的话就按“0”尽快反应,如果不一样则不需要反应。60张图片统一处理成8 cm × 11 cm的彩色图片,每张图片水平视角大约为 6°,垂直视角大约为 8.4 °(或水平视角为 8.4°,垂直视角为 6°)。
2.4 ERP记录和数据处理
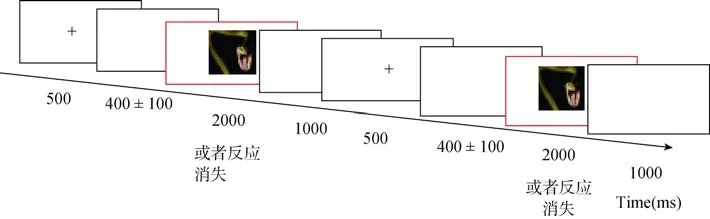
图1 实验流程示意图
采用德国 BP公司的脑电记录仪 Active Two BioSemi 系统(BioSemi,Amsterdam,Netherlands),记录50导电极的信号。以左耳乳突为参考电极点,闲置原有的另一只参考电极,取TP8电极连于右乳突,形成单极导联。离线(Offline)分析时以置于右乳突的有效电极进行再参考,即从各导联信号中减去1 /2该参考电极所记录的信号。在左眼四周常规放置4个电极记录垂直和水平眼电。头皮与电极之间的阻抗小于 5 kΩ,记录连续 EEG,频带宽度为0.1~20 Hz,每导采样频率为 500 Hz,通过 Brain Vision Analyzer software(BrainProducts)进行离线分析。分析时程是从每张图片呈现前的100 ms持续到图片呈现的1000 ms。本文主要分析N1、N2、P3和慢波(Slow wave,SW),N1、N2选取Fz、FCz和Cz等3个头皮前部电极进行分析;P3和SW选取CPz和Pz等2个头皮顶部电极进行分析。由于N1和 N2这两个负成分起始和结束的时间都很短,并且中间还夹着一个P1成分,为了尽可能排除P1成分的干扰,因此分别取这两个成分峰值左右 10 ms的平均波幅进行统计分析。最后对N1(120~140 ms)和N2(230~250 ms) 两个成分分别进行2(焦虑水平:低焦虑,高焦虑)×2(图片:中性图片,恐惧图片)×3(电极点:Fz、FCz 和 Cz),以及对 P3(300~500 ms)和SW(500~1000 ms)两个成分分别进行2(焦虑水平:低焦虑,高焦虑)×2(图片:中性图片,恐惧图片)×2(电极点:CPz、和Pz)三因素重复测量方差分析,其中电极点和图片为被试内因素,焦虑水平为被试间因素。
3 结果
图2表示在 Fz、FCz、Cz等 3个电极点上的ERP波形,其N1、N2成分清晰可见;图3表示CPz、Pz上的P3和SW成分。

图2 高低焦虑个体加工中性图片和恐惧图片时在N1和N2两个成分上的差异
3.1 N1
三因素重复测量方差分析结果发现,焦虑水平的主效应显著,F
(1,32)=11.94,p
<0.01,即高特质焦虑的个体诱发的 N1平均波幅(−5.08 µV)显著地比低焦虑的个体(−2.27 µV)更大。电极点的主效应显著,F
(2,64)=19.28,p
<0.01,Fz点的平均波幅(−4.01 µV)与 FCz 的平均波幅(−3.95 µV)差异不显著,但是均显著地大于 Cz点的平均波幅(−3.05µV)。没有发现显著的图片类型的主效应,但焦虑水平×图片×电极点的交互作用显著,F
(1,32)=3.40,p
<0.05。进一步分析发现焦虑水平×图片的交互作用显著,F
(1,32)=6.88,p
<0.05,简单效应检验结果如图4所示,在 Cz和 FCz两个电极点,发现高特质焦虑的个体组的图片主效应显著[Cz:F
(1,32)=6.93,p
<0.05,FCz:F
(1,32)=5.22,p
<0.05],具体为恐惧图片比中性图片诱发了一个更加大的 N1,而低特质焦虑的组中则发现两种图片诱发的N1没有显著的差异,说明了高焦虑的个体进行情绪加工时,在120~140 ms时就开始区分中性图片和恐惧图片,而低特质焦虑的个体在120~140 ms时还没能区分这两种图片。3.2 N2
三因素重复测量方差分析结果发现,焦虑水平的主效应显著,F
(1,32)=4.54,p
<0.05,即低特质焦虑的个体诱发的 N2波幅(−5.72 µV)显著大于高焦虑的个体(−2.22 µV)。图片类型的主效应也显著,F
(1,32)=4.83,p
<0.05,具体为中性图片(−4.50µV)比恐惧图片(−3.44 µV)诱发了一个更大的 N2。电极点的主效应显著,F
(2,64)=22.80,p
<0.01,具体为 Fz 点波幅最大(−5.23 µV),FCz(−4.20 µV)居中,Cz 点(−2.48 µV)最小,两两之间差异均显著。焦虑水平、图片和电极点三个因素构成的交互作用都不显著,这些结果说明在230~250 ms时,高、低特质焦虑个体都能区分中性图片和恐惧图片。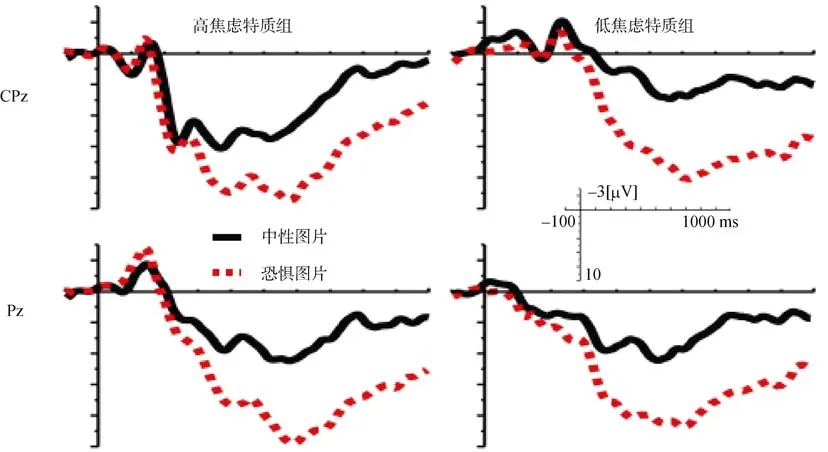
图3 高低焦虑个体加工中性图片和恐惧图片时P3、SW波形

图4 在N1和N2两个成分上图片和焦虑水平的交互作用
3.3 P3
三因素重复测量方差分析结果,发现显著的图片类型的主效应,F
(1,32)=69.5,p
<0.01,具体为恐惧图片(6.61 µV)比中性图片(3.21 µV)诱发了一个更加大的 P3;还发现电极点的主效应显著,F
(1,32)=38.69,p
<0.01,具体表现为Pz点的平均波幅(5.94 µV)显著地比 CPz 的波幅(3.87 µV)更大(如图3)。焦虑水平的主效应以及焦虑水平、图片和电极点三个因素构成的交互作用都不显著。3.4 SW
三因素重复测量方差分析结果发现,只有图片类型的主效应显著,F
(1,32)=84.12,p
<0.01,具体为恐惧图片(6.93 µV)比中性图片(2.51 µV)诱发了一个更加大的慢波。电极点的主效应、焦虑水平的主效应、以及焦虑水平、图片和电极点三个因素构成的交互作用都不显著。3.5 特质焦虑得分与ERPs的关系
本研究以Fz、FCz和Cz三点记录到的个体加工恐惧图片时N1、N2以及CPz、Pz记录到的P3、SW 的波幅的平均值作为因变量,以特质焦虑的得分(STAI-T总分)作为自变量进行回归,同时为了排除性别、年龄的影响,把性别、年龄也纳入回归分析中,进行多元逐步回归分析。控制性别和年龄的贡献后,结果发现STAI-T总分能够预测N2的波幅,却不能预测N1、P3和SW的波幅。最终的回归方程为 Y=−15.34+0.29 X,F
=6.97,p
<0.05,R
=0.18(其中,Y=N2波幅值,X=特质焦虑的得分),说明特质焦虑的得分和加工恐惧图片所诱发的N2的波幅值之间的呈正相关。因为 N2为负成分,波幅值与N2波幅本身成反比,即波幅值越小说明N2波形越往负方向偏移,N2的波幅越大。所以,特质焦虑的得分和加工恐惧图片所诱发的N2的波幅之间的呈负相关,也就是说,特质焦虑水平越低的个体,由恐惧图片诱发的N2越大。4 讨论
大量的行为研究一致发现了焦虑个体存在着对负性刺激的选择性注意偏向的特点(Bar-Haim et al.,2007),Holmes等人(2008)的ERP研究也发现以面孔为刺激材料时焦虑个体的 P1、P2效应,揭示了高特质焦虑者加工情绪面孔时早期注意偏向的特点,但他们未关注 N1、N2等负成分,不能不说是一种缺憾。本研究拟着重比较高低特质焦虑者的N1、N2、P3等ERP成分,以探讨对复杂情景中的威胁信息的选择性注意偏向发生的时间进程及神经机制。我们的研究结果发现,高特质焦虑者早在刺激后120~140 ms时就开始区分恐惧图片和中性图片,表现为恐惧图片比中性图片诱发出一个更大的N1,而低焦虑的个体却没有发现这种N1效应。Mogg和 Bradley(2002)提出了关于焦虑水平影响选择性注意偏向的认知-动机模型(cognitivemotivational model),该模型认为,对威胁信息的注意由效价评价系统所决定,这个系统负责刺激的最初的无意识的评价,信息从这个系统输出这一认知过程中,会受到动机因素例如焦虑水平的影响。高特质焦虑会提高对威胁信息的敏感性,使个体对威胁信息投入更多的注意资源(Mogg &Bradley,1998)。我们的结果表明,高特质焦虑者由于对威胁信息更加敏感,使其对复杂情景中的威胁信息进行情绪加工时投入更多的注意资源,表现出了对威胁信息的知觉迅速(开始于 120~140 ms)并且需要消耗更多的注意资源,因此支持了 Mogg的认知-动机模型。
此外,不管特质焦虑水平的高低,都发现中性图片诱发的N2显著大于恐惧图片诱发的N2,说明了在 230~250 ms,低特质焦虑个体也开始能够区分恐惧图片和中性图片。图片的主效应持续到了晚期的P3和SW,在高低特质焦虑两组人都发现,恐惧图片诱发的P3和SW显著大于中性图片诱发的P3和SW。这些结果说明了,不管焦虑水平的高低,个体能够在230~250 ms便开始区分出中性图片和恐惧图片,并且在情绪加工后期给予恐惧图片更多的注意资源。尽管高特质焦虑的个体这两个成分的平均波幅都稍微高于低特质焦虑的个体,但是差异不显著。这可能是因为P3和SW反映了刺激意义的改变(Hajcak &Nieuwenhuis,2006),本研究的实验范式要求被试识别即可,并没有要求被试对图片意义的深入加工,所以两组被试在这两个成分上没有显著差异是可以理解的。另外还可能是因为 P3和SW与动机密切相关(Schupp et al.,2000;Schupp,Junghöfer,Weike,&Hamm 2004),而从生物进化论的角度上来说,威胁性刺激例如恐怖的动物,与人类的生存紧密相关(Öhman,1993),人类需要对危险信号迅速地知觉并投入更多地注意资源,这种对威胁相关信息的注意加工偏向应该是一种适应性的表现。本实验中的两组被试尽管特质焦虑的水平上存在显著差异,但是都属于正常人,都同样能够迅速地识别恐惧情绪,因此尽管在本研究中,这两个成分存在差异,但是却没有达到显著水平,后续的研究可以比较正常人和焦虑症患者情绪加工的特点,以便更好地说明问题。
回归分析的结果发现,特质焦虑得分越高的个体,N2波幅越小。前人关于焦虑与执行控制能力的研究也发现了类似的结果(Eldar &Bar-Haim,2010),他们认为,N2成分与抑制执行功能密切相关(Folstein &van Petten,2008;Nieuwenhuis,Yeung,van den Wildenberg,&Ridderinkhof,2003;van Veen&Carter,2002),高焦虑个体N2变小,反映了个体的抑制执行功能可能受损。所以我们认为N2反映了个体调用抑制执行功能的有效性,焦虑水平越低的个体,越能够有效地抑制情绪信息,因此表现出越大的N2。同样,个体能够有效地抑制对中性图片的情绪的加工,因此表现出更加大的 N2,而恐惧图片由于占用了个体比较多的注意资源,因此削弱了对其情绪的加工的抑制,所以表现出比较小的N2,进而在方差分析的结果中表现为中性图片诱发的 N2显著大于恐惧图片,并且低特质焦虑的个体相对于高特质焦虑的个体诱发更加大的 N2。这些结果都说明了高特质焦虑个体对恐惧图片进行情绪加工时不能有效地调用抑制执行功能。
综上所述,本研究结果说明低特质焦虑的个体在 230~250 ms开始区分恐惧图片和中性图片,并且迅速地对调用抑制执行能力对恐惧图片进行进一步的加工,使其对恐惧图片的情绪加工保持在适应性的范围内。而高特质焦虑的个体在情绪加工早期120~140 ms时就开始区分恐惧图片和中性图片,分配过多的注意资源,并且不能有效地对情绪进行抑制加工。因此,这种对威胁信息的过分警惕,且不能很好地抑制,很可能是高特质焦虑个体选择性注意偏向重要原因。尽管本研究提供的结果能够为我们理解高特质焦虑个体发展成临床焦虑障碍患者提供一定的理论依据(Li et al.,2005;Li et al.,2007),但是本研究也存在着一些不足,例如本研究采用 block设计以及没有收集状态焦虑水平,可能会干扰试验结果,但是由于这些对高、低特质焦虑的两组被试的干扰是一致的,所以根据心理实验设计中的减法原则,比较高低特质焦虑的两组被试时,这些无关因素对本研究造成的干扰是可以被排除的。尽管如此,进一步研究需要对状态焦虑进行控制,以及补充假随机呈现刺激时数据作为比较,以排除无关变量的影响。
总之,本研究采用 ERP的技术考察了高低特质焦虑的个体在进行情绪加工时的注意偏向的神经机制。结果发现高特质焦虑者相对于低特质焦虑者诱发了较大N1,并且恐惧图片诱发的N1比中性图片的 N1更大,说明了高特质焦虑者早在120~140 ms时就对恐惧图片分配了较多的注意资源;而低特质焦虑者在 230~250 ms时开始区分恐惧图片和中性图片,并且相对高特质焦虑者诱发了更大的 N2,并且特质焦虑得分越高的个体,N2波幅越小,说明了高特质焦虑者的抑制执行功能可能受损。
Bai,L.,Ma,H.,Huang,Y.X.,&Luo,Y.J.(2005).The development of native Chinese affective picture system-a pretest in 46 college students.Chinese Mental Health Journal,19
(11),719-722.[白露,马慧,黄宇霞,罗跃嘉.(2005).中国情绪图片系统的编制——在46名中国大学生中的试用.中国心理卫生杂志,19
(11),719-722.]Bar-Haim,Y.,Lamy,D.,&Glickman,S.(2005).Attentional bias in anxiety:Abehavioral and ERP study.Brain and Cognition,59
(1),11-22.Bar-Haim,Y.,Lamy,D.,Pergamin,L.,Bakermans-Kranenburg,M.J.,&van IJzendoorn,M.H.(2007).Threat-related attentional bias in anxious and nonanxious individuals:Ameta-analytic study.Psychological Bulletin,133
(1),1-24.Bradley,B.P.,Mogg,K.,White,J.,Groom,C.,&Bono,J.(1999).Attentional bias for emotional faces in generalized anxiety disorder.British Journal of Clinical Psychology,38
(3),267-278.Dennis,T.A.,&Chen,C.C.(2009).Trait anxiety and conflict monitoring following threat: An ERP study.Psychophysiology,46
(1),122-131.Eldar,S.,&Bar-Haim,Y.(2010).Neural plasticity in response to attention training in anxiety.Psychological Medicine,40
(4),667-677.Folstein,J.R.,&van Petten,C.(2008).Influence of cognitive control and mismatch on the N2 component of the ERP:Areview.Psychophysiology,45
(1),152-170.Foti,D.,&Hajcak,G.(2008).Deconstructing reappraisal:Descriptions preceding arousing pictures modulate the subsequent neural response.Journal of Cognitive Neuroscience,20
(6),977-988.Foti,D.,Hajcak,G.,&Dien,J.(2009).Differentiating neural responses to emotional pictures: Evidence from temporal-spatial PCA.Psychophysiology,46
(3),521-530.Gao,P.C.,&Huang,M.E.(2008).Attentional bias of high trait anxious individuals.Acta Psychologica Sinica,40
(3),307-318.[高鹏程,黄敏儿.(2008).高焦虑特质的注意偏向特点.心理学报,40
(3),307-318.]Gilboa-Schechtman,E.,Foa,E.B.,&Amir,N.(1999).Attentional biases for facial expressions in social phobia:The face-in-the-crowd paradigm.Cognition &Emotion,13
(3),305-318.Hajcak,G.,&Nieuwenhuis,S.(2006).Reappraisal modulates the electrocortical response to unpleasant pictures.Cognitive,Affective,&Behavioral Neuroscience, 6
,291-297.Hajcak,G.,MacNamara,A.,&Olvet,D.M.(2010).Event-related potentials,emotion,and emotion regulation:An integrative review.Developmental Neuropsychology,35
(2),129-155.Hajcak,G.,Weinberg,A.,MacNamara,A.,&Foti,D.(2012).ERPs and the Study of Emotion
.In S.J.Luck &E.S.Kappenman(Eds.),Oxford handbook of event-related potential components
(pp.441-474).New York:Oxford University Press.Holmes,A.,Nielsen,M.K.,&Green,S.(2008).Effects of anxiety on the processing of fearful and happy faces:An event-related potential study.Biological Psychology,77
(2),159-173.Kessler,R.C.,McGonagle,K.A.,Zhao,S.,Nelson,C.B.,Hughes,M.,Eshleman,S.,&Kendler,K.S.(1994).Lifetime and 12-month prevalence of DSM-III-R psychiatric disorders in the United States:Results from the National Comorbidity Survey.Archives of General Psychiatry,51
(1),8-19.Lang,P.J.,Bradley,M.M.,&Cuthbert,B.N.(1999).International affective picture system(IAPS)
:Technical manual and affective ratings
.Gainesville,FL:The Center for Research in Psychophysiology,University of Florida.Lang,P.J.,Davis,M.,&Öhman,A.(2000).Fear and anxiety:Animal models and human cognitive psychophysiology.Journal of Affective Aisorders,61
(3),137-159.Li,X.Y.,Li,X.B.,&Luo,Y.J.(2005).Anxiety and attentional bias for threat:An event-related potential study.Neuroreport,16
(13),1501-1505.Li,X.Y.,Wang,M.,Poliakoff,E.,&Luo,Y.J.(2007).Attention to threat in high and low trait-anxious individuals:Astudy using extremely threating pictorial cues.Perceptual and Motor Skills,104
(3c),1097-1106.Luo,W.,Feng,W.,He,W.,Wang,N.-Y.,&Luo,Y.-J.(2010).Three stages of facial expression processing:ERP study with rapid serial visual presentation.NeuroImage,49
,1857-1867.MacNamara,A.,&Hajcak,G.(2009).Anxiety and spatial attention moderate the electrocortical response to aversive pictures.Neuropsychologia,47
(13),2975-2980.Mangun,G.R.,&Hillyard,S.A.(1990).Electrophysiological studies of visual selective attention in humans.In A.B.Scheibel &A.Wechsler(Eds.),The neurobiological foundations of higher cognitive function
(pp.271-295).New York:Guilford Press.McNally,R.J.(2002).Anxiety and its disorders:The nature and treatment of anxiety and panic,2nd ed.American Journal of Psychiatry,159
(8),1453-1453.Mogg,K.,&Bradley,B.P.(1998).Acognitive-motivational analysis of anxiety.Behaviour Research and Therapy,36
(9),809-848.Mogg,K.,&Bradley,B.P.(2002).Selective orienting of attention to masked threat faces in social anxiety.Behaviour Research and Therapy,40
(12),1403-1414.Nieuwenhuis,S.,Yeung,N.,van den Wildenberg,W.,&Ridderinkhof,K.R.(2003).Electrophysiological correlates of anterior cingulate function in a go/no-go task:Effects of response conflict and trial type frequency.Cognitive,Affective,&Behavioral Neuroscience,3
(1),17-26.Schupp,H.T.,Cuthbert,B.N.,Bradley,M.M.,Cacioppo,J.T.,Ito,T.,&Lang,P.J.(2000).Affective picture processing:The late positive potential is modulated by motivational relevance.Psychophysiology,37
,257-261.Schupp,H.T.,Junghöfer,M.,Weike,A.I.,&Hamm,A.O.(2004).The selective processing of briefly presented affective pictures:An ERP analysis.Psychophysiology,41
,441-449.Shek,D.T.L.(1993).The Chinese version of the state-trait anxiety inventory:Its relationship to different measures of psychological well-being.Journal of Clinical Psychology,49
(3),349-358.Spielberger,C.D.,Gorsuch,R.L.,Lushene,R.,Vagg,P.R.,&Jacobs,G.A.(1983).Manual for the state-trait anxiety inventory
.[Press].Palo Alto,CA:Consulting Psychologist.Öhman,A.(1993).Fear and anxiety as emotional phenomena:Clinical phenomenology,evolutionary perspectives,and information-processing mechanisms.In:M.Lewis,&J.M.Haviland(Eds.),Handbook of emotions
(pp.511-536).New York:Guilford Press.van Veen,V.,&Carter,C.S.(2002).The timing of action-monitoring processes in the anterior cingulate cortex.Journal of Cognitive Neuroscience,14
(4),593-602.Weinberg,A.,&Hajcak,G.(2010).Beyond good and evil:The time-course of neural activity elicited by specific picture content.Emotion,10
(6),767-782.