不同红锥种源幼林光合特性日变化测定与分析
2013-01-03李志辉朱积余
蒋 燚 ,李志辉 ,朱积余 ,姜 英
(1.中南林业科技大学,湖南 长沙410004;2.广西林业科学研究院,广西 南宁530002;3.广西优良用材林资源培育重点实验室,广西 南宁530002;4.国家林业局中南速生材繁育实验室,广西 南宁530002)
不同红锥种源幼林光合特性日变化测定与分析
蒋 燚1,2,3,4,李志辉1,朱积余2,3,4,姜 英2,3,4
(1.中南林业科技大学,湖南 长沙410004;2.广西林业科学研究院,广西 南宁530002;3.广西优良用材林资源培育重点实验室,广西 南宁530002;4.国家林业局中南速生材繁育实验室,广西 南宁530002)
采用英国生产的LCPRO+全自动便携式植物光合测定仪,以10个种源红锥幼林叶片为研究对象,进行光合特性测定和比较分析。结果表明:不同红锥种源净光合速率、蒸腾速率和气孔导度日变化均呈“双峰”曲线,在上午10:00(主峰值)和下午17:30(次峰值)表现出2个峰值;其净光合速率在2个峰值时间段中间有明显的“午休”现象,其产生原因是自身的气孔限制因子;红锥种源的胞间CO2浓度为不规则“W”型,以清早为最高,不同种源的最小值出现的时间有所不同;10个种源的生长性状指标与净光合速率呈显著正相关性(其中树高、材积与净光合速率呈极显著性正相关,胸径与净光合速率呈显著性正相关)。在以木材收获量为主要目标的人工林经营中,应选择材积量和净光合速率最大的博白和浦北红锥种源。
红锥;净光合速率;蒸腾速率;气孔导度;胞间CO2浓度;种源
作为我国南亚热带季风常绿阔叶林的优势种或建群种[1-2]的红锥Castanopsis hystrix A.DC.,是华南地区重要的乡土阔叶珍贵用材和高效多用途树种。自上世纪80年代初开始进行该树种资源调查研究以来,在引种驯化、丰产栽培、育苗技术[3]等方面已开展了近30年的相关研究。近年来,在红锥的木材性质[4-5]、微生物菌肥肥效[6]、遗传多样性[7]等方面也开展了相关的研究,其生长快、材质优、适应广、效益高等优良特性,已得到科研和生产部门的认同。近10年来,珍贵树种的发展得到了进一步的重视,随着种植面积的不断增加,对珍贵树种的良种要求也在不断提升,因此,优良种源的选择显得尤为重要。选择优良种源其目的是能有效地利用种源的优良性状[8],很多学者对一些树种种源的光合作用进行一定的研究,大部分树种的种源不同其光合特性差异很明显,表现在季节变化、日变化等差异上,但亦有少数树种不同种源间的光合速率没有显著差异,这与树种本身特性相关。植物光合作用本身是一个复杂的生理过程,光合能力的强弱在一定程度上取决于物种的遗传特性[9],日均净光合速率(Pn)能够反映植物光合能力的大小[10]。本实验对6.5年生红锥幼林10个种源的进行了净光合速率日变化测定,分析不同种源红锥幼林的光合特性与其生长指标的相关关系,有助于揭示红锥各种源的生长规律及其差异性, 对红锥种源试验、良种选育、适地适树、造林技术及抚育管理、创造高生产力的人工林生态系统提供有关依据。
1 材料与方法
1.1 供试材料来源
参试材料来自广西苍梧县林科所6.5年生的10个红锥种源,10个种源分别来自广西的浦北、博白、容县、苍梧、凭祥、宁明、东兰和广东的陆河、信宜、高州10个地点(表1),种源试验林面积18 hm2,设置3次重复,每种源为1小区种120株。试验采用随即区组设计,株行距2 m×3 m,带状整地,规格50 cm×50 cm×40 cm。穴施基肥磷肥0.25 kg,营养杯苗造林,种植后当年和第二年各抚育一次,并每次追肥复合肥150 g。

表1 红锥种源来源地概况Table 1 Profile of provenances of C. hystrix
1.2 光合特性测定
测定仪器:英国生产的LCPRO+全自动便携式植物光合测定仪,选择标准叶室。
测定时期:8月21日~25日。
测定植株和叶样选择:10个参试红锥种源均选择生长健壮且无病虫害的3个待测植株,选择树冠中部向阳的第2年生的第4~6轮叶片(从顶端数),进行光合作用日变化测定;测定叶片确定后,做好挂牌标记并记录,每个种源分别从3个待测株上选取5~6片叶。
测定指标:净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、 胞间CO2浓度(Ci)等。
光合日变化测定方法:选择晴朗无云天气,测定时间段为7:00~19:00每隔1.5 h测定1次,3次重复,分析数据取平均值;测定前调试好仪器,测定时利用光合测定系统的控温装置调节叶室温度在24~30℃之间,待叶片适应叶室环境3 min,数据稳定后记录。
1.3 生长指标测定
测定时间:2008年10月20日~22日。
测定指标:树高、胸径、冠幅。
测定方法:全测试验林全部林木,树高采用测高杆直接量取,以m记录并保留一位小数;树高用林木测树尺测量,以cm记录保留一位小数;冠幅取树冠边缘按上下、左右用皮尺量取,以m记录并保留一位小数。
1.4 计算统计与分析
运用Excel 2003和 SPSS15.0 统计分析软件系统对数据进行分析处理。
2 结果与分析
2.1 净光合速率(Pn)日变化
10个红锥种源叶片的光合速率日变化曲线(如图1),其形状均为一种不对称的“M”形,在上午10:00(主峰值)和下午17:30(次峰值)表现出2个峰值,2个峰值时间段中间有明显的“午休”现象;在10个种源中主峰值以浦北种源和博白种源的峰值最大,分别为 13.53 和 12.04 μmol·m-2s-1,以东兰种源的峰值(6.49 μmol·m-2s-1)最小,仅为浦北和博白峰值的48.0%和53.9%。按Pn的主峰值从大到小排序,依次为浦北>博白>陆河>宁明>苍梧>容县>高州>信宜>凭祥>东兰;按Pn的日平均值从大到小排序,依次为浦北>博白>陆河>苍梧>宁明>容县>高州>信宜>东兰>凭祥,分别为6.80、5.86、4.45、4.40、4.25、4.14、3.63、3.59、3.24 和 3.18 μmol·m-2s-1。

图1 净光合速率Pn日变化Fig.1 Daily changes of Pn
植物的光合作用是一个复杂的生理过程,叶片光合效率与光照强度、气温、CO2浓度、水分等因子有关[11],在同科植物卷斗青冈、细叶青冈、青冈栎[12]、赤皮青冈[13]、白栎和藜蒴[14]等树种均有相同之处。本次试验的结果,从上午7:00~10:00期间,红锥种源光合速率随光合有效辐射和气温的升高而不断升高,出现第一个峰值;10:00~14:00期间,光合速率随光合有效辐射和气温的继续升高反而下降,表明在此期间气温升高、蒸腾作用加强、叶片水分减少,导致气孔部分关闭,造成光合作用原料即胞间CO2浓度降低,使光合速率下降,出现较为明显地光合“午休”现象。
2.2 蒸腾速率(Tr)日变化
红锥不同地理种源蒸腾速率日变化与净光合速率有相似的变化趋势(图2),其日变异规律是:早晚蒸腾速率低,上午10:00达主峰,之后下降,14:30为低谷,然后上升,至17:30达次峰,之后再下降。按主峰值从大到小的排序为浦北>博白>陆河>苍梧>高州>容县>宁明>凭祥>信宜>东兰,其中浦北为4.32 mmol·m-2s-1,东兰为 3.28 mmol·m-2s-1。
2.3 气孔导度(Gs)日变化

图2 蒸腾速率Tr日变化Fig.2 Daily changes of Tr
气孔是植物叶片与外界进行气体交换的通道,气孔导度表示的是气孔张开的程度,影响光合作用、呼吸作用及蒸腾作用,气孔导度的变化对植物水分状况及CO2同化有着重要意义。气孔导度减小,CO2进入叶肉细胞内的阻力增加,从而导致光合速率下降,同时气孔阻力的增加也减少叶片水分散失,在一定程度上减轻干旱胁迫对光合器官的伤害。可见气孔导度是影响植物光合速率的主要因子之一[15-16]。由图3可知,10个红锥种源气孔导度日进程规律是:其性状均为“双峰”型;在10:00和17:30有2个峰值,10:00时峰值明显高于17:30时峰值;所有在全天中以傍晚时气孔导度值最低。10个红锥种源之间差异不大,其中以浦北(0.225 mmol·m-2s-1) 的峰值最大,东兰(0.173 mmol·m-2s-1)峰值最小。
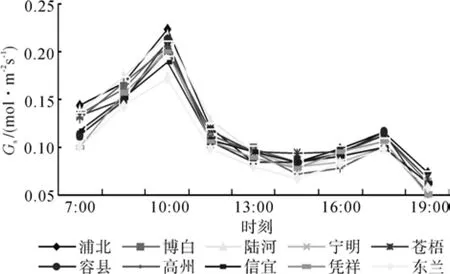
图3 气孔导度Gs日变化Fig.3 Daily changes of Gs
2.4 胞间 CO2浓度(Ci)日变化
由图4可知,10个红锥家系的胞间CO2浓度均呈不规则“w”形状,峰值均出现在7:00、16:00和19:00(信宜种源除外),其中以7:00的值最高。而最小值出现的时间各有不同,其中宁明、苍梧、高州、信宜和凭祥的值于11:30达到全天的最小值,依次为219.0、149.8、188.5、170.0、222.0 和 157.0 μmol·mol-1;浦北和陆河最小值出现在 13:00,为 209.5 和 205.0 μmol·mol-1;博白、容县和东兰于17:30达到全天最小,分别为184.5、187.0 和 150.5 μmol·mol-1。

图4 胞间CO2浓度Ci日变化Fig.4 Daily changes of Ci
2.5 不同种源红锥叶片光合生理特性参数的相关分析
取10个种源红锥日变化各光合特征参数的平均值进行相关分析(表2),可以看出,10个种源Pn与Tr均呈极显著正相关,浦北和苍梧2个种源的Pn与Gs呈显著正相关,剩余8个种源的Pn与Gs呈极显著正相关,Pn与Ci的相关性不显著。

表2 10个种源红锥净光合速率与其他光合生理特性参数的相关分析†Table 2 Correlation analysis of ten provenances of C. hystrix net photosynthetic rate and other physiological parameters
2.6 红锥生长指标与净光合速率的相关性分析
通过对红锥生长指标与净光合速率的相关性分析(表3),可以看出,红锥种源的Pn与其树高H和材积V呈极显著性正相关,与胸径D呈显著性正相关,表明净光合速率越高红锥生长越快。

表3 红锥生长指标及净光合速率的相关性分析†Table 3 Correlation analysis between C. hystrix growth index and net photosynthetic rate
3 结论与讨论
(1)林木本身的遗传特性是影响其光合能力的主要因素,也受环境因子的影响,如叶绿素含量、叶片厚度、叶片成熟程度、叶片有效光辐射、温度、湿度、土壤含水量等,从而使林木的光合速率及其相关因子在森林群区之内或之间存在很大的差异[17-20]。参与本试验10个红锥种源虽来自不同的10个种源点,鉴于红锥自身的特点,从表1也可看出,10个种源点气候环境较为相似,同时,所采集进行光合测定的叶片源于气候土壤等条件基本一致的同一试验地点,10个种源光合日进程均是一条不对称的“双峰”曲线, 均在上午10: 00和下午17:30 各有一个峰值,出现了明显且持续时间较长的光合“午休”现象,可以说明同一树种的红锥有相似的遗传特性;但在净光合速率峰值上有差异,其中净光合速率以浦北和博白最大,东兰最小,这也说明,10个红锥种源由于种源来源地环境影响致使种源间在有净光合速率一定的差距。本结论中净光合速率变化趋势与王蕾在优质红锥遗传多样性分析中[21]中5月所测结论一致,但光合速率大小及排序有所不同,这正好印证宋宏伟研究不同种源黄连木苗光合特性[22]指出的不同种源的净光合速率和蒸腾速率随测定时间的不同而不同的结论。
(2)产生光合“午休”的原因,研究者[23-24]认为形成光合“午休”因素有2个,即气孔限制因素和非气孔限制因素。前者是由于强光、高温、低湿度、低CO2浓度等环境因子引起气孔部分关闭,造成Ci降低而引起Pn下降;后者是由于叶肉细胞自身羧化酶活性(羧化阻力增加)降低导致Pn下降,此时Ci增高。两因素同时存在时,Ci变化倾向于优势因素。因此,在10个红锥种源的净光合速率午后降低时,此时气温、光照达到当日的最高值,而Gs和Ci均降低(图3和图4),这说明10个种源红锥的Pn午间降低出现光合“午休”主要是由气孔限制因素引起的。
(3)在本试验中各种源的光合速率与主要受温度驱动的维度梯度之间没有显著的关系,种源地自然环境条件较为复杂(表1),很难断定确切的主导环境因子,关于导致这10个红锥种质产生较大光合差异的其它因子还需要进一步研究。但这种光合能力之间的种源差异是能在子代中得以表现的遗传型适应的结果[25-26],所以在进行品种选育时要选择Pn较高的种源红锥作为试验材料,这样会有效地利用光照资源,产生更多的有机物,从而获得高产的木材,对于参试的10个红锥种源净光合速率最大为浦北和博白种源,建议再今后的生产中尽量采用这2个红锥种源。
[1] 《广西森林》编辑委员会编著.广西森林[M].北京:中国林业出版社,2001:234-239.
[2] 《中国森林》编辑委员会编著.中国森林[M].北京:中国林业出版社,2000:570-1574.
[3] 蒋 燚,朱积余,张泽尧,等.红锥种源多点育苗试验研究[J].广西林业科学,2005,34(4):196-199.
[4] 陈桂丹,唐贤明,覃引弯,等.人工林红锥树皮率及树皮纤维尺寸的研究[J].中南林业科技大学学报,2012,32(6):163-167.
[5] 蒋 燚,李志辉,朱积余,等.红锥家系木材密度等物理性状遗传和变异性分析[J].中南林业科技大学学报,2012,32(11):9-13,20.
[6] 胡彩颜,康丽华,江业根,等.三个乡土树种苗期微生物菌肥施肥效应的研究[J].中南林业科技大学学报,2011,31(12):18-24.
[7] 杨 峰,李志辉,蒋 燚,等.红锥优良家系ISSR遗传多样性分析[J].中南林业科技大学学报,2012,32(6):123-127.
[8] 马德华编著.植物育种的数量遗传学基础[M].南京:江苏科学技术出版社,1982.
[9] 何文兴,易津,李洪梅.根茎禾草乳熟期净光合速率日变化的比较研究[J].应用生态学报,2004,15(2):205-209.
[10] 傅松玲,黄成林,曹恒生,等.黄山松更新特性与光因子关系的研究[J].应用生态学报,2000,11(6):801-804.
[11] 潘瑞炽,董愚得.植物生理学[M].北京:高等教育出版社,1999.
[12] 吴丽君,李志辉,邹 峰. 水分胁迫对珍贵树种赤皮青冈幼苗生理特性的影响[J].中南林业科技大学学报,2012,32(7):10-13.
[13] 李志辉,卢兆银,杨 艳,等.松栎混交林中马尾松与白栎光合作用的日变化[J].中南林业科技大学学报,2007,27(1):78-80.
[14] 黄志宏,周光益,邱治军.南岭森林3种常见树种光合-蒸腾作用特性[J].中南林业科技大学学报,2009,29(2):18-24.
[15] 陈存及,邱尔发,梁一池,等.毛竹不同种源光合特性研究[J].林业科学,2001,37(6):16-19.
[16] 刘建福.澳州果叶片光合速率和叶绿素荧光参数日变化[J].西南农业大学学报, 2006,28(2):271-273.
[17] Oquist G. Stress and adaption in photosynthesis[C]//Douglas R H,Moan J. Lightinbiology and medicine. New York: Plenum Press,1988,433.
[18] Dur L Z, Goldstein G. Photosynthesis, photoinhibition, and nitrogenuse efficiency in native and invasive tree ferns in Hawaii[J].Oeco logia,2001,126(3):345-354.
[19] 褚延广,苏晓华,黄秦军,等.欧洲黑杨基因资源光合生理特征与生长的关系[J].林业科学,2010,46(7):77-83.
[20] 杜克兵,许 林,涂炳坤,等.淹水胁迫对2种杨树1年生苗叶片超微结构和光合特性的影响[J].林业科学,2010,46(6):58-65.
[21] 王 蕾.优质红锥遗传多样性分析[D].厦门:厦门大学,2006
[22] 宋宏伟,于海燕,卢绍辉,等.不同种源黄连木苗光合特性的研究[J].林业科学研究, 2009,22(3):454-457.
[23] 许大全.光合作用“午睡”现象的生态、生理与生化[J].植物生理学通讯,1990,26(6):52.
[24] Tenhunen J D. Factors in fluencing carbon fixation and water useby mediterranean chlorophyll shrubs during summer drought[J]. Oecologia,1990,82:381-393.
[25] 曹福亮,蔡金峰,汪贵斌,等.2010.淹水胁迫对乌桕生长及光合作用的影响[J].林业科学,46(10):57-61.
[26] 赵晓焱,王传宽,霍 宏.2008.兴安落叶松光合能力及相关因子的种源差异[J].生态学报,28(8):3799-3806.
Diurnal variations of photosynthetic characteristics of different provenances Castanopsis hystrix A.DC. young plantations
JIANG Yi1,2,3,4, LI Zhi-hui1, ZHU Ji-yu2,3,4, JIANG Ying2,3,4
(1.Central South University of Forestry and Technology, Changsha 410004, Hunan, China; 2. Guangxi Academy of Forestry, Nanning 530002, Guangxi, China; 3.Guangxi Key Laboratory of Superior Timber Trees Resource Cultivation, Nanning 530002, Guangxi, China;4.Key Laboratory of Central South Fast-growing Timber Cultivation, Nanning 530002, Guangxi, China)
With LCPRO+ Automatic Portable Plant Photosynthesis Meter made in British, by taking 10 different provenances of Castanopsis hystrix young leaves as the studying objects, the photosynthetic characteristics and comparative analysis have been investigated. The results show that the net photosynthetic rate, transpiration rate and diurnal variations of stomatal conductance of different C. hystrix provenances appeared a bimodal curve, being at 10:00 in the morning and 17:30 in the afternoon; there was an obvious “noon break” phenomenon between the 2 peak time’s interspace, the reason is their own stomatal limitation factors; the intercellular CO2concentration of C. hystrix provenances appeared inregular W-type, the highest concentration of CO2appeared in the morning, and the minimum values of different provenances appeared in different time quantum; the growth traits and rate of net photosynthesis of 10 different provenances were positive correlation each other, the tree height and wood volume showed an extremely significant positive correlation to net photosynthesis rate, the diameter at breast height and net photosynthetic rate showed a significant positive correlation. It is suggested that Bobai and Pubei C. hystrix provenances that have maximum amount of volume and net photosynthetic rate should be chosen for the plantation management which takes timber yield as the main target.
Castanopsis hystrix A.DC.; transpiration rate; stomatal conductance; intercellular CO2concentration; provenance
S718.155
A
1673-923X(2013)06-0043-05
2013-02-27
广西科学研究与技术开发计划课题“红锥良种选育与丰产栽培模式研究”(桂科攻10100012-3)
蒋 燚(1968-),男,广西灌阳人,教授级高级工程师,博士研究生,主要从事森林培育研究;E-mail:jy68@163.com
李志辉(1957-),男,湖南安化人,教授,博士生导师,主要从事森林培育教学和研究;E-mail:lzh1957@126.com
[本文编校:欧阳钦]
