10个秋菊品种的光合特性及净光合速率与部分生理生态因子的相关性分析
2012-12-31李永华张开明于红芳
李永华,张开明,于红芳
在园林绿化实践中,园林植物的选择和应用不但要考虑其美学需要,更要考虑其光合生理特性。众多学者对园林植物的光合特性进行了研究[1-2],并探讨了园林植物对环境的适应性及其光合作用机制,为园林植物的合理配置及科学管理提供了重要的理论依据和指导。
菊花〔Dendranthema morifolium(Ramat.)Tvzel.〕为菊科(Compositae)菊属〔Dendranthema(DC.)Des Moul.〕多年生草本植物,是重要的园林观赏花卉之一,以秋菊盆栽观赏为主。目前,国内学者对菊花的起源、品种分类、花期调控及栽培生理等[3-6]方面均进行了较为深入的研究,但对其光合特性方面的研究较少,且主要集中在逆境胁迫下菊花光合生理指标的变化等[7-8]方面,对不同秋菊品种的光合特性则缺少系统的比较分析。
作者以常见的秋菊5个早花品种及5个晚花品种为研究对象,对它们的光合参数进行了测定,以揭示其光合生理特征;并分析了不同品种的光合生理特性差异及其与环境因子之间的相互关系,以了解不同秋菊品种的光合作用基本规律,为秋菊的栽培管理和合理配置提供理论依据和技术指导。
1 材料和方法
1.1 材料
供试材料为秋菊早花品种‘早粉盘’、‘铜雀春深’、‘檀香狮子’、‘太平的小鼓’和‘日出东方’以及晚花品种‘关东新侠’、‘云龙凤舞’、‘绿牡丹’、‘星光灿烂’和‘日本黄’,均由河南省开封市禹王台公园提供。
于2009年5月在河南农业大学实验基地进行扦插育苗,7月上盆。花盆选择透气性好的瓦盆,栽培基质为堆肥土、园土、草木灰和细沙的混合基质,按照体积比2∶2∶1∶1的比例混合均匀,每盆1株,采用常规管理。
1.2 方法
于2009年9月至10月,分别选取生长旺盛且长势一致的健壮植株(株高30 cm、约有20片叶),采用CIRAS-2便携式光合测定系统(英国PP-SYSTEM公司)测定顶芽下第3至第4片完全展开的功能叶的各项光合指标值。测定期间实验地午间最大光照强度约1 500μmol·m-2·s-1,最高气温35℃,大气相对湿度约30%。每品种选取3株,各指标分别重复测定5次,结果取平均值。
1.2.1 净光合速率的光响应曲线绘制 于晴天上午9:00至10:00分别在光合有效辐射强度(PAR)为0、50、100、150、200、250、300、400、500、600、700、800、900、1 000、1 100、1 200、1 300、1 400和1 500μmol·m-2· s-1的条件下测定叶片净光合速率(Pn)。将Pn和PAR进行二元回归分析,并计算各品种的光补偿点(LCP)、光饱和点(LSP)、最大净光合速率(Pnmax)及相关性,Pn和PAR的直线回归方程的初始斜率即为光合作用的表观量子效率(AQY)。测定时,设定光合测定系统的叶室温度为(25±1)℃,CO2浓度为(380±5)μmol·mol-1,大气相对湿度为70%。
1.2.2 净光合速率的CO2响应曲线绘制 于晴天上午9:00至10:00分别在胞间CO2浓度(Ci)为0、50、100、150、200、250、300、400、500、600、700、800、900、1 000、1 100、1 200、1 300、1 400和1 500 μmol·m-2·s-1的条件下测定叶片Pn。将Pn和Ci进行二元回归分析,并计算各品种的 CO2补偿点(CCP)、CO2饱和点(CSP)、最大净光合速率(Pnmax)及相关性,Pn和Ci的直线回归方程的初始斜率即为光合作用的羧化效率(CE)。测定时,设定光合测定系统的叶室光照强度为900μmol·m-2· s-1,温度为(25±1)℃,大气相对湿度为70%。
1.2.3 光合特性参数日变化的测定 于晴天的6:00至18:00分别选取各植株同一部位叶片测定Pn,每隔2 h测定1次。同时,记录叶片的蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)、光合有效辐射强度(PAR)、大气CO2浓度(Ca)、大气温度(Ta)和大气相对湿度(RH)。其中,PAR、Ca、Ta和RH以外界条件为准;为了消除时间上的误差,重复测定时各品种随机排序。
1.3 数据处理
采用Excel 2003软件进行数据处理和制表,并采用SPSS 17.0软件(中文版)进行数据的相关性分析、线性回归分析及其他统计分析。
2 结果和分析
2.1 叶片光响应参数的比较
净光合速率(Pn)的光响应曲线能够反映植物叶片的光合速率随光合有效辐射强度(PAR)改变的变化规律。由供试10个秋菊品种Pn的光响应曲线(图略)可知:各品种的Pn变化趋势均呈相似的二次曲线。在PAR低于200μmol·m-2·s-1的条件下,随PAR的增加,Pn迅速增大;随PAR的继续增加,Pn上升幅度减小,达到最大值后渐渐趋于平衡。以PAR为自变量x、Pn为因变量y进行线性回归分析,获得的各秋菊品种的回归方程较相似(表1)。
由表1可看出:10个秋菊品种的光饱和点(LSP)存在极显著差异,多数品种的光补偿点(LCP)、最大净光合速率(Pnmax)和表观量子效率(AQY)也存在极显著差异。10个秋菊品种的LSP为962.51~1 077.53μmol·m-2·s-1,LCP为92.83~167.37 μmol·m-2·s-1。可见,秋菊属喜光植物,对光照的需求较高。品种‘日本黄’的LSP和LCP均最低,分别为962.51和92.83μmol·m-2·s-1,表明其耐阴性较强,但对弱光的利用能力不强;品种‘云龙凤舞’的LSP和LCP也均较低,但极显著高于品种‘日本黄’的LSP和LCP;而品种‘早粉盘’及‘檀香狮子’的LSP和LCP均较高,其中,品种‘早粉盘’的LSP和LCP分别为1 077.53和141.17μmol·m-2·s-1,品种‘檀香狮子’的LSP和LCP分别为1 035.44和134.28μmol·m-2·s-1,表明二者的耐阳性较强;品种‘太平的小鼓’和‘铜雀春深’具有较高的LSP和较低的LCP,说明二者对光照的适应能力较强;品种‘日出东方’、‘关东新侠’、‘绿牡丹’及‘星光灿烂’均具有较高的LCP和较低的LSP,说明它们对光照的适应范围较窄;品种‘云龙凤舞’的LSP、LCP和AQY也较低,表明其耐阴能力也较强,但对弱光的利用能力则较弱。
表1 10个秋菊品种叶片光响应参数的比较(±SE)1)Table 1 Com parison of light response parameters of leaves of ten cultivars of autumn chrysanthemum〔Dendranthema morifolium(Ramat.) Tvzel.〕(±SE)1)

表1 10个秋菊品种叶片光响应参数的比较(±SE)1)Table 1 Com parison of light response parameters of leaves of ten cultivars of autumn chrysanthemum〔Dendranthema morifolium(Ramat.) Tvzel.〕(±SE)1)
1)同列中不同的大写和小写字母分别表示差异极显著(P<0.01)或显著(P<0.05)Different capitals and small letters in the same column indicate extremely significant difference(P<0.01)or significant difference(P<0.05),respectively.LSP:光饱和点Light saturation point;LCP:光补偿点Light compensation point;Pnmax:最大净光合速率Maximum net photosynthetic rate;AQY:表观量子效率Apparent quantum yield.
品种Cultivar LSP/μmol·m-2·s-1 LCP/μmol·m-2·s-1 Pnmax/μmol·m-2·s-1 AQY 回归方程Regression equation早粉盘Zaofenpan 1 077.53±40.72Aa 141.17±5.56Bb 17.53±0.75Aa 0.044 5±0.002 0Aa y=0.044 5x -6.081铜雀春深Tongquechunshen 1 045.34±39.81Bb 120.19±4.98Ee 17.10±0.69Bb 0.031 9±0.001 5Gh y=0.031 9x-4.175檀香狮子Tanxiangshizi 1 035.44±33.14Cc 134.28±5.66Cc 16.22±0.73Cc 0.042 1±0.002 0Ccd y=0.042 1x-5.597太平的小鼓Taipingdexiaogu 1 030.13±35.53Dd 103.72±4.10Ff 17.16±0.58Bb 0.039 2±0.001 8De y=0.039 2x-4.163日出东方Richudongfang 1 022.52±40.14Ee 167.37±5.98Aa 14.62±0.66Gg 0.043 2±0.002 0Bb y=0.043 2x-6.780关东新侠Guandongxinxia 1 017.53±31.53Ff 141.53±3.98Bb 15.35±0.61Ee 0.042 4±0.001 9Cc y=0.042 4x-5.724云龙凤舞Yunlongfengwu 1 010.24±39.42Gg 120.23±3.76Ee 15.83±0.64Dd 0.038 6±0.001 7Ef y=0.038 6x-4.703绿牡丹Lümudan 1 000.35±32.14Hh 128.61±5.10Dd 15.19±0.59EFf 0.039 2±0.001 8De y=0.039 2x-4.960星光灿烂Xingguangcanlan 985.42±29.93Ii 128.04±4.62Dd 14.69±0.57Gg 0.041 9±0.002 1Cd y=0.041 9x-5.219日本黄Ribenhuang 962.51±30.21Jj 92.83±3.92Gg 15.13±0.65Ff 0.036 2±0.001 5Fg y=0.036 2x -3.436
2.2 叶片CO2响应参数的比较
供试的10个秋菊品种叶片的CO2饱和点(CSP)、CO2补偿点(CCP)、最大净光合速率(Pnmax)以及净光合速率(Pn)与胞间CO2浓度(Ci)的回归方程见表2。各品种Pn的CO2响应曲线(图略)的变化趋势基本相同;以Ci为自变量x、净光合速率为因变量y进行线性回归分析,获得的10个品种的回归方程(表2)也较为相似。在Ci低于400μmol·mol-1的条件下,随Ci的增加,Pn呈直线增长趋势;随Ci的继续增加,Pn增幅减小,并逐渐达到最大值,之后趋于平稳。当Ci为300~800μmol·mol-1时,10个秋菊品种的Pn为6.75~24.43μmol·m-2·s-1,可见,Ci增加可显著提高菊花的Pn值。
从表2可见:10个品种的CSP差异极显著,多数品种的CCP及Pnmax有极显著差异。10个品种的CCP为77.62~133.16μmol·mol-1,表明它们为典型的C3植物。10个品种的 CSP为1 060.46~1 485.48 μmol·mol-1,其中,品种‘关东新侠’和‘绿牡丹’的CSP分别为1 485.48和1 456.25μmol· mol-1,极显著高于其他品种;品种‘檀香狮子’和‘早粉盘’的CSP则较低,极显著低于其他品种。而品种‘星光灿烂’和‘绿牡丹’的CCP分别为77.62和87.83 μmol·mol-1,极显著低于其他品种,表明这2个品种对低浓度胞间CO2的利用能力较强。
表2 10个秋菊品种叶片CO2响应参数的比较(±SE)1)Table 2 Comparison of CO2 response parameters of leaves of ten cultivars of autumn chrysanthemum〔Dendranthema morifolium(Ramat.) Tvzel.〕(±SE)1)

表2 10个秋菊品种叶片CO2响应参数的比较(±SE)1)Table 2 Comparison of CO2 response parameters of leaves of ten cultivars of autumn chrysanthemum〔Dendranthema morifolium(Ramat.) Tvzel.〕(±SE)1)
1)同列中不同的大写和小写字母分别表示差异极显著(P<0.01)或显著(P<0.05)Different capitals and small letters in the same column indicate extremely significant difference(P<0.01)or significant difference(P<0.05),respectively.CSP:CO2饱和点CO2 saturation point;CCP:CO2补偿点CO2 compensation point;Pnmax:最大净光合速率Maximum net photosynthetic rate;CE:羧化效率Carboxylation efficiency.
品种Cultivar CSP/μmol·mol-1 CCP/μmol·mol-1 Pnmax/μmol·m-2·s-1 CE 回归方程Regression equation早粉盘Zaofenpan 1 100.46±41.24Ii 100.31±4.02De 19.99±0.76Dd 0.042 3±0.002 0Bb y=0.042 3x -4.217铜雀春深Tongquechunshen 1 285.31±47.43Ff 100.15±4.06De 28.07±1.36Aa 0.052 6±0.002 5Aa y=0.052 6x-5.244檀香狮子Tanxiangshizi 1 060.46±40.41Jj 133.16±5.46Aa 19.18±0.61Fg 0.040 3±0.001 9Cc y=0.040 3x-5.237太平的小鼓Taipingdexiaogu 1 212.54±42.54Gg 107.85±4.54Cd 24.43±0.95Bb 0.038 1±0.001 6Dd y=0.038 1x-4.257日出东方Richudongfang 1 132.53±43.32Hh 102.75±3.88De 21.21±0.69Cc 0.040 4±0.002 0Cc y=0.040 4x-4.311关东新侠Guandongxinxia 1 485.48±51.43Aa 111.21±4.21Cc 18.87±0.81Ee 0.034 3±0.001 6Ff y=0.034 3x-3.677云龙凤舞Yunlongfengwu 1 361.11±50.31Dd 122.91±4.93Bb 13.82±0.58Hj 0.022 5±0.001 3Ii y=0.022 5x-2.839绿牡丹Lümudan 1 456.25±54.14Bb 87.83±3.21Fg 14.98±0.63Gh 0.027 2±0.001 2Hh y=0.027 2x-2.473星光灿烂Xingguangcanlan 1 400.36±48.32Cc 77.62±3.98Gh 17.49±0.66Ff 0.028 8±0.001 5Ee y=0.028 8x-3.008日本黄Ribenhuang 1 305.28±49.84Ee 93.64±3.69Ef 14.67±0.42Gi 0.028 3±0.001 1Gg y=0.028 3x -2.687
2.3 叶片光合参数日变化的比较
供试的10个秋菊品种叶片光合参数的日变化见表3。由表3可以看出:在6:00至10:00,10个品种的净光合速率(Pn)均呈快速上升的趋势,于10:00左右达到峰值(11~19μmol·m-2·s-1);在10:00至14:00,Pn迅速下降,至14:00左右降到低谷值(1~6μmol·m-2·s-1);然后Pn又缓慢上升,在16:00前后达到次峰值;之后Pn开始下降,18:00左右降到最低值(1~3μmol·m-2·s-1)。10个秋菊品种光合参数的日变化均表现出“上升-下降-再上升-再下降”的趋势,说明这些品种的光合作用存在明显的“午休”现象。然而,10个秋菊品种的光合能力有一定差异,品种‘早粉盘’和‘铜雀春深’对午前上升的光照强度较敏感,Pn快速增加,能充分利用上午的有利环境条件进行光合作用;而品种‘檀香狮子’则从“午休”状态中恢复较快,可以利用午后较长的时间进行相对较强的光合作用。
由表3还可见:10个品种的气孔导度(Gs)和蒸腾速率(Tr)的日变化趋势与Pn相似,区别在于Gs的峰值出现在10:00左右,与Pn的最大值同步出现;而Tr的峰值则出现在12:00左右;Gs与Tr的低谷值均出现在14:00左右;Gs与Tr的次峰值均出现在16:00前后。胞间CO2浓度(Ci)的日变化趋势与Pn基本相反,Ci在6:00、14:00及18:00左右较高,在10:00及16:00出现低谷值,2个Ci低谷值出现的时间与Pn的2个峰值一致。
表3 10个秋菊品种叶片光合参数日变化的比较(±SE)1)Table 3 Com parison of daily changes of photosynthetic parameters of leaves of ten cultivars of autumn chrysanthemum〔Dendranthem morifolium(Ramat.)Tvzel.〕(±SE)1)
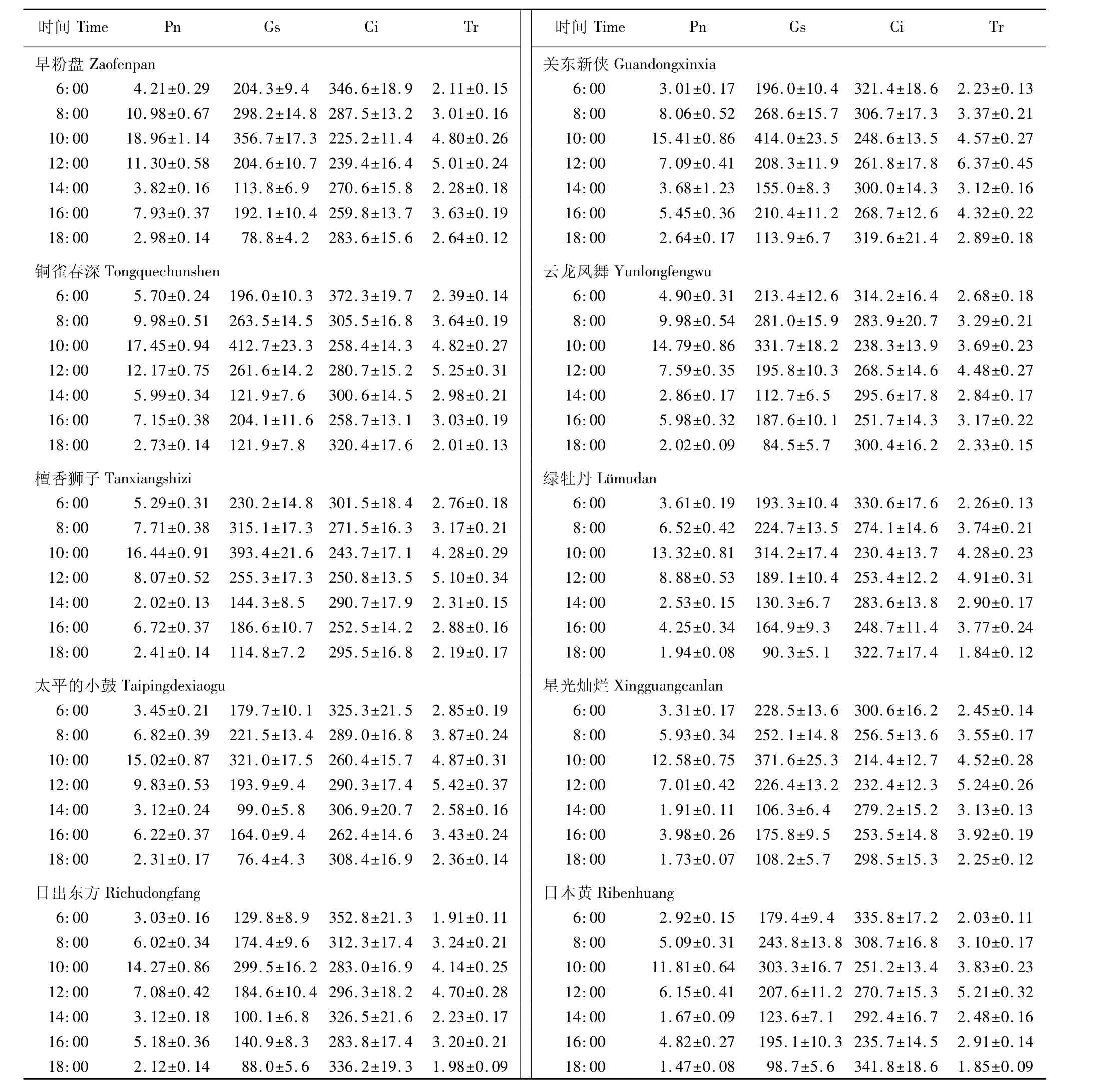
表3 10个秋菊品种叶片光合参数日变化的比较(±SE)1)Table 3 Com parison of daily changes of photosynthetic parameters of leaves of ten cultivars of autumn chrysanthemum〔Dendranthem morifolium(Ramat.)Tvzel.〕(±SE)1)
a 1)Pn:净光合速率Net photosynthetic rate;Gs:气孔导度 Stomatal conductance;Ci:胞间CO2浓度Intercellular CO2 concentration;Tr:蒸腾速率Transpiration rate.
时间Time Pn Gs Ci Tr 时间Time Pn Gs Ci Tr早粉盘Zaofenpan 关东新侠Guandongxinxia 6:00 4.21±0.29 204.3±9.4 346.6±18.9 2.11±0.15 6:00 3.01±0.17 196.0±10.4 321.4±18.6 2.23±0.13 8:00 10.98±0.67 298.2±14.8 287.5±13.2 3.01±0.16 8:00 8.06±0.52 268.6±15.7 306.7±17.3 3.37±0.21 10:00 18.96±1.14 356.7±17.3 225.2±11.4 4.80±0.26 10:00 15.41±0.86 414.0±23.5 248.6±13.5 4.57±0.27 12:00 11.30±0.58 204.6±10.7 239.4±16.4 5.01±0.24 12:00 7.09±0.41 208.3±11.9 261.8±17.8 6.37±0.45 14:00 3.82±0.16 113.8±6.9 270.6±15.8 2.28±0.18 14:00 3.68±1.23 155.0±8.3 300.0±14.3 3.12±0.16 16:00 7.93±0.37 192.1±10.4 259.8±13.7 3.63±0.19 16:00 5.45±0.36 210.4±11.2 268.7±12.6 4.32±0.22 18:00 2.98±0.14 78.8±4.2 283.6±15.6 2.64±0.12 18:00 2.64±0.17 113.9±6.7 319.6±21.4 2.89±0.18铜雀春深Tongquechunshen 云龙凤舞Yunlongfengwu 6:00 5.70±0.24 196.0±10.3 372.3±19.7 2.39±0.14 6:00 4.90±0.31 213.4±12.6 314.2±16.4 2.68±0.18 8:00 9.98±0.51 263.5±14.5 305.5±16.8 3.64±0.19 8:00 9.98±0.54 281.0±15.9 283.9±20.7 3.29±0.21 10:00 17.45±0.94 412.7±23.3 258.4±14.3 4.82±0.27 10:00 14.79±0.86 331.7±18.2 238.3±13.9 3.69±0.23 12:00 12.17±0.75 261.6±14.2 280.7±15.2 5.25±0.31 12:00 7.59±0.35 195.8±10.3 268.5±14.6 4.48±0.27 14:00 5.99±0.34 121.9±7.6 300.6±14.5 2.98±0.21 14:00 2.86±0.17 112.7±6.5 295.6±17.8 2.84±0.17 16:00 7.15±0.38 204.1±11.6 258.7±13.1 3.03±0.19 16:00 5.98±0.32 187.6±10.1 251.7±14.3 3.17±0.22 18:00 2.73±0.14 121.9±7.8 320.4±17.6 2.01±0.13 18:00 2.02±0.09 84.5±5.7 300.4±16.2 2.33±0.15檀香狮子Tanxiangshizi 绿牡丹Lümudan 6:00 5.29±0.31 230.2±14.8 301.5±18.4 2.76±0.18 6:00 3.61±0.19 193.3±10.4 330.6±17.6 2.26±0.13 8:00 7.71±0.38 315.1±17.3 271.5±16.3 3.17±0.21 8:00 6.52±0.42 224.7±13.5 274.1±14.6 3.74±0.21 10:00 16.44±0.91 393.4±21.6 243.7±17.1 4.28±0.29 10:00 13.32±0.81 314.2±17.4 230.4±13.7 4.28±0.23 12:00 8.07±0.52 255.3±17.3 250.8±13.5 5.10±0.34 12:00 8.88±0.53 189.1±10.4 253.4±12.2 4.91±0.31 14:00 2.02±0.13 144.3±8.5 290.7±17.9 2.31±0.15 14:00 2.53±0.15 130.3±6.7 283.6±13.8 2.90±0.17 16:00 6.72±0.37 186.6±10.7 252.5±14.2 2.88±0.16 16:00 4.25±0.34 164.9±9.3 248.7±11.4 3.77±0.24 18:00 2.41±0.14 114.8±7.2 295.5±16.8 2.19±0.17 18:00 1.94±0.08 90.3±5.1 322.7±17.4 1.84±0.12太平的小鼓Taipingdexiaogu 星光灿烂Xingguangcanlan 6:00 3.45±0.21 179.7±10.1 325.3±21.5 2.85±0.19 6:00 3.31±0.17 228.5±13.6 300.6±16.2 2.45±0.14 8:00 6.82±0.39 221.5±13.4 289.0±16.8 3.87±0.24 8:00 5.93±0.34 252.1±14.8 256.5±13.6 3.55±0.17 10:00 15.02±0.87 321.0±17.5 260.4±15.7 4.87±0.31 10:00 12.58±0.75 371.6±25.3 214.4±12.7 4.52±0.28 12:00 9.83±0.53 193.9±9.4 290.3±17.4 5.42±0.37 12:00 7.01±0.42 226.4±13.2 232.4±12.3 5.24±0.26 14:00 3.12±0.24 99.0±5.8 306.9±20.7 2.58±0.16 14:00 1.91±0.11 106.3±6.4 279.2±15.2 3.13±0.13 16:00 6.22±0.37 164.0±9.4 262.4±14.6 3.43±0.24 16:00 3.98±0.26 175.8±9.5 253.5±14.8 3.92±0.19 18:00 2.31±0.17 76.4±4.3 308.4±16.9 2.36±0.14 18:00 1.73±0.07 108.2±5.7 298.5±15.3 2.25±0.12日出东方Richudongfang 日本黄Ribenhuang 6:00 3.03±0.16 129.8±8.9 352.8±21.3 1.91±0.11 6:00 2.92±0.15 179.4±9.4 335.8±17.2 2.03±0.11 8:00 6.02±0.34 174.4±9.6 312.3±17.4 3.24±0.21 8:00 5.09±0.31 243.8±13.8 308.7±16.8 3.10±0.17 10:00 14.27±0.86 299.5±16.2 283.0±16.9 4.14±0.25 10:00 11.81±0.64 303.3±16.7 251.2±13.4 3.83±0.23 12:00 7.08±0.42 184.6±10.4 296.3±18.2 4.70±0.28 12:00 6.15±0.41 207.6±11.2 270.7±15.3 5.21±0.32 14:00 3.12±0.18 100.1±6.8 326.5±21.6 2.23±0.17 14:00 1.67±0.09 123.6±7.1 292.4±16.7 2.48±0.16 16:00 5.18±0.36 140.9±8.3 283.8±17.4 3.20±0.21 16:00 4.82±0.27 195.1±10.3 235.7±14.5 2.91±0.14 18:00 2.12±0.14 88.0±5.6 336.2±19.3 1.98±0.09 18:00 1.47±0.08 98.7±5.6 341.8±18.6 1.85±0.09
2.4 净光合速率与主要生理生态因子的相关性分析
供试10个秋菊品种叶片的净光合速率(Pn)与气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)、光合有效辐射强度(PAR)、大气温度(Ta)、大气相对湿度(RH)和大气CO2浓度(Ca)的相关系数见表4。由表4可见:10个品种的Pn与Gs的相关系数为0.892~0.987,均表现为极显著的正相关关系;Pn与Tr的相关系数为0.652~0.906,其中品种‘早粉盘’、‘铜雀春深’及‘太平的小鼓’的Pn与Tr极显著正相关,品种‘檀香狮子’、‘日出东方’、‘绿牡丹’及‘星光灿烂’的Pn与Tr显著正相关,其他品种的Pn与Tr呈不显著的正相关关系;10个品种Pn与Ci的相关系数为-0.898~-0.759,其中品种‘早粉盘’、‘铜雀春深’、‘檀香狮子’、‘星光灿烂’及‘日本黄’的Pn与Ci极显著负相关,其他品种的Pn与Ci均呈显著的负相关关系。此外,10个品种的Pn与PAR和Ta正相关,与RH和Ca负相关,但相关性均不显著。
实验结果表明:10个秋菊品种叶片的净光合速率主要受气孔导度和胞间CO2浓度的影响,与蒸腾速率、光合有效辐射强度、大气温度、大气相对湿度和大气CO2浓度也有一定的相关性。
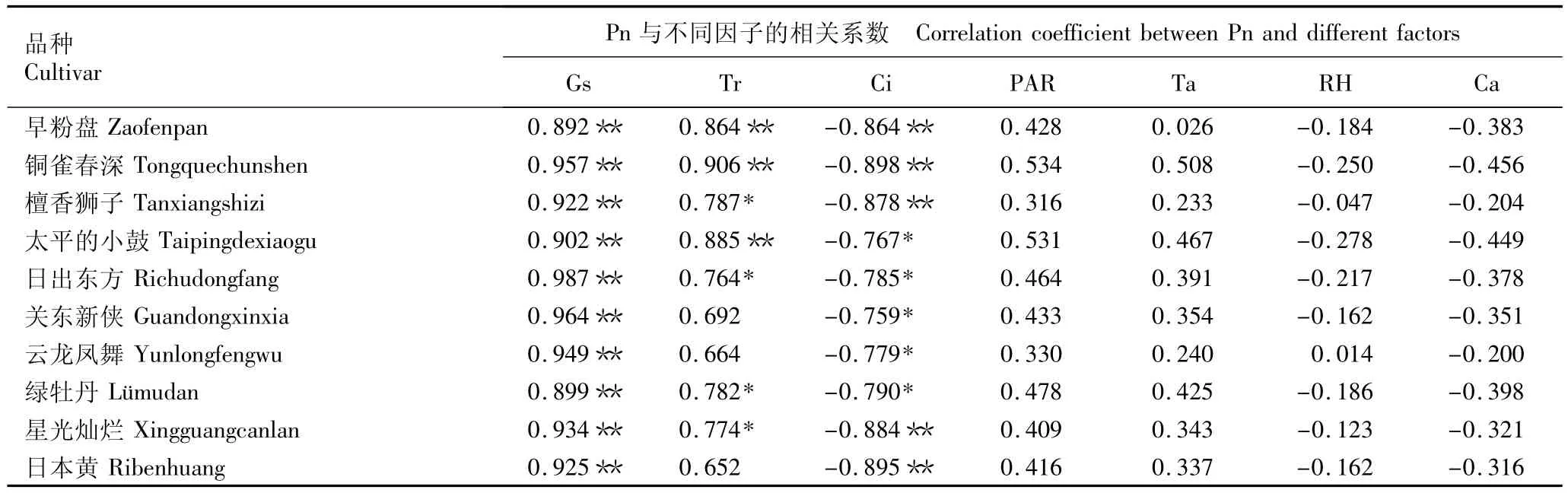
表4 10个秋菊品种叶片的净光合速率(Pn)与主要生理生态因子的相关系数1)Table 4 Correlation coefficients between net photosynthetic rate(Pn)and main physio-ecological factors in leaves of ten cultivars of autumn chrysanthemum〔Dendranthema morifolium(Ramat.)Tvzel.〕1)
3 讨论和结论
植物叶片光合能力的强弱在一定程度上取决于物种的遗传特性[1-2,9]。光饱和点和光补偿点可用来衡量植物对强光或弱光的利用能力,光饱和点较高的种类对强光环境较为适应,而较高的光补偿点可导致产物消耗量增加[9-10]。植物的表观量子效率越低,其对光能的利用能力越弱。供试10个秋菊品种的光饱和点为962.51~1 077.53μmol·m-2·s-1,光补偿点为92.83~167.37μmol·m-2·s-1,表明这些秋菊品种均属于喜光植物。秋菊不同品种对光照条件的利用能力不同,各品种之间差异显著。早花品种中,品种‘早粉盘’及‘檀香狮子’的光饱和点、光补偿点和表观量子效率均较高,表明这2个品种在强光环境下能够有效进行光合作用,但产物消耗量较多;品种‘太平的小鼓’和‘铜雀春深’具有较高的光饱和点以及较低的光补偿点和表观量子效率,表明这2个品种对光照条件的适应范围较大,适宜在光照较强的环境下栽培;品种‘日出东方’具有较高的光补偿点和表观量子效率以及较低的光饱和点,表明其适应的光照条件范围较窄,但利用光能的能力较强且能在相对较弱的光照环境中生长。晚花品种中,品种‘关东新侠’、‘绿牡丹’和‘星光灿烂’具有较高的光补偿点和表观量子效率以及较低的光饱和点,能够在相对较弱的光照条件下利用光能,可以在树阴下等光照条件相对较弱的环境中进行栽植;品种‘日本黄’和‘云龙凤舞’的光饱和点、光补偿点和表观量子效率均较低,表明这2个品种具有一定的耐阴性,但其利用弱光的能力相对较弱。
CO2是光合作用的重要原料,其浓度直接影响植物的光合作用。CO2浓度升高,一方面增加了CO2对Rubisco酶结合位点的竞争能力,进而提高羧化效率;另一方面可抑制光呼吸作用,进而提高光合效率[11]。供试的10个秋菊品种的CO2补偿点(77.62~133.16 μmol· mol-1)远大于一般常见的C3植物 (30~70 μmol·mol-1),说明这些秋菊品种均为典型的C3植物;它们的羧化效率为0.022 5~0.052 6,均明显低于常见的 C3植物 (0.06~0.12)。与美丽异木棉(Chorisia speciosa St.)[12]相比,供试的10个秋菊品种的CO2补偿点和CO2饱和点(1 060.46~1 485.48 μmol·mol-1)均较高,表明秋菊对低浓度CO2的利用能力不强。但是,不同秋菊品种对CO2的利用能力差异显著,其中品种‘绿牡丹’、‘星光灿烂’及‘日本黄’的CO2补偿点均较低,CO2饱和点均较高,表明这3个品种对低浓度及高浓度CO2的利用能力均较强。作者的研究结果显示:当胞间CO2浓度为300~800 μmol·mol-1时,10个秋菊品种的净光合速率为6.75~24.43μmol·m-2·s-1,可见,适当增加CO2浓度可显著提高秋菊的净光合速率。在实际生产中,可以通过增施CO2肥料等措施来提高菊花的光合速率。
供试的10个秋菊品种净光合速率(Pn)的日变化趋势与许多植物[13-18]相似,呈典型的双峰型曲线,首峰均出现在10:00左右,次峰均出现在16:00左右,并有明显的光合“午休”现象;早花品种的Pn普遍高于晚花品种,在10:00,早花品种的Pn为14.27~18.96μmol·m-2·s-1,而晚花品种的Pn为11.81~15.41μmol·m-2·s-1。10个秋菊品种的气孔导度和蒸腾速率的日变化规律与净光合速率相似,也均呈双峰型曲线;而胞间CO2浓度的日变化趋势基本上与净光合速率相反。
相关性分析结果表明:供试的10个秋菊品种的净光合速率与气孔导度呈极显著正相关,与胞间CO2浓度呈显著或极显著的负相关,与光合有效辐射强度和大气温度呈不显著正相关,与大气相对湿度和大气CO2浓度呈不显著负相关;除品种‘关东新侠’、‘云龙凤舞’和‘日本黄’外,其他7个品种的净光合速率均与蒸腾速率有显著或极显著的正相关关系。上述结果表明:秋菊叶片的光合作用受气孔导度、胞间CO2浓度、蒸腾速率和光合有效辐射强度等生理生态因子的综合影响,其中气孔导度和胞间CO2浓度的影响较大。其他生理生态因子与菊花光合作用的关系及其作用机制有待进一步研究。
综合上述研究结果,建议在秋菊的生产中,在强光、高温及低湿的夏季对菊花进行适当遮阳和喷灌,以降低空气温度、增加空气相对湿度,从而减少植株的水分蒸腾消耗,避免由于强光照射而引起的叶片光合结构的破坏,进而减弱光合作用的“午休”现象,提高菊花的光合作用生产力。
[1]张友焱,周泽福,党宏忠,等.毛乌素沙地油蒿不同生长期的枝条水势和叶片气体交换特性[J].植物资源与环境学报,2010,19(3):17-22.
[2]于晓霞,阮成江.曼陀罗光合特性研究[J].植物资源与环境学报,2011,20(1):40-45.
[3]戴思兰,王文奎,黄家平.菊属系统学及菊花起源的研究进展[J].北京林业大学学报,2002,24(5/6):230-234.
[4]张树林.菊花品种分类的研究[J].园艺学报,1965,4(1):35-46.
[5]杨 娜,郭维明,陈发棣,等.光周期对秋菊品种‘神马’花芽分化和开花的影响[J].园艺学报,2007,34(4):965-972.
[6]许 瑛,陈发棣.菊花8个品种的低温半致死温度及其抗寒适应性[J].园艺学报,2008,35(4):559-564.
[7]孙宪芝,郑成淑,王秀峰.高温胁迫对切花菊‘神马’光合作用与叶绿素荧光的影响[J].应用生态学报,2008,19(10): 2149-2154.
[8]贾思振,房伟民,陈发棣,等.高温下5个夏菊品种开花特性、叶片组织结构与光合特性的比较[J].南京农业大学学报,2009,32(3):151-156.
[9]靳甜甜,刘国华,胡婵娟,等.黄土高原常见造林树种光合蒸腾特征[J].生态学报,2008,28(11):5758-5765.
[10]傅松玲,黄成林.黄山松更新特性与光因子关系的研究[J].应用生态学报,2000,11(6):801-804.
[11]蒋跃林,张庆国,杨书运,等.28种园林植物对大气CO2浓度增加的生理生态反应[J].植物资源与环境学报,2006,15 (2):1-6.
[12]吴吉林,李永华,叶庆生.美丽异木棉光合特性的研究[J].园艺学报,2005,32(6):1061-1064.
[13]靳忠英,彭正松,李育明,等.半夏的光合特性[J].作物学报,2006,32(10):1542-1548.
[14]侯小改.4个牡丹品种光合特性的比较研究[J].河南农业大学学报,2007,41(5):527-530.
[15]吴彦琼,胡玉佳.外来植物南美蟛蜞菊、裂叶牵牛和五爪金龙的光合特性[J].生态学报,2004,24(10):2334-2339.
[16]黄成林,傅松玲,梁淑云,等.五种攀缘植物光合作用与光因子关系的初步研究[J].应用生态学报,2004,15(7):1131-1134.
[17]陶 俊,陈 鹏.银杏光合特性的研究[J].园艺学报,1999,26(3):157-160.
[18]杨江山,常永义,种培芳.樱桃不同节位叶片光合特性与解剖特征比较研究[J].果树学报,2005,22(4):323-326.
