转Bar基因抗除草剂稻谷对小鼠致敏性的研究
2012-11-22孙艳波段妍慧富丽娜颜亨梅
孙艳波,黄 毅,段妍慧,富丽娜,刘 金,颜亨梅
(湖南师范大学生命科学学院,中国 长沙 410081)
大米虽然被认为是低致敏食品,并且是唯一可以免于过敏性试验的谷物,但是仍有对大米过敏的人存在,而采用了转基因技术的稻谷的潜在致敏性更是转基因食品安全评价的焦点之一[1-2].随着食品安全问题日益受到国家政府的重视,相关国际组织均建立了致敏性评估程序.目前转基因食品致敏性评价一般首先根据FAO/WHO[3]和Codex Alimentarius Commission[4]建议的致敏性评估决策树途径,无论表达蛋白是否来源于已知过敏原,评价时都将其序列与已知过敏原的抗原决定簇比对,分析序列同源性;并利用生物信息学工具来预测蛋白质的三维结构,高级结构的构建将有助于阐明蛋白质在分子水平上的效应机制[5].之后再利用动物致敏模型进行研究,用转基因作物食品直接喂养实验动物,以观察转基因植物/食品的食用安全性.国内外许多实验室相关研究多为28 d营养学评价实验或90 d亚慢性实验,其得出的实验结果均为无显著性差异[6-8].本实验考虑到小肠是主要的消化、吸收器官,对肠道免疫屏障进行研究对致敏性的评价有重要意义[9].因此,本研究选用供试小鼠肠道屏障系统中肠粘液sIgA水平和释放到血清中的DAO水平,加之外周血血清中IgE检测3项指标作为过敏性检测指标,综合判断取食转抗除草剂Bar基因稻谷后的小鼠是否出现致敏性反应或病变.
1 材料和方法
1.1 材料
稻谷:实验品种Bar68-1(农基安审字(2006)第060号)为中国科学院亚热带农业生态研究所培育抗草丁膦早籼稻;对照品种D68为Bar68-1的非转基因亲本.
饲料:基础饲料[生产许可证号:SCXK(湘)2009-0009],并参照国际上公认的符合小鼠生长发育繁殖要求的AIN-93G[10]的经典饲料配方(全价营养饲料)而重新配比制作的饲料,转基因、常规水稻的掺入质量分数为60%(保证蛋白质质量分数不低于18%,脂肪质量分数不低于3.0%,以确保小鼠正常的生长发育需要,具体配方见表1).
实验动物:SPF级昆明小鼠60只,体重18~24 g[生产许可证号:SCXK(湘)2009-0004].
主要试剂:小鼠sIgA、DAO、IgE ELISA定量测定试剂盒(武汉USCN life公司).
主要仪器:高速冷冻离心机(TCL-16A),酶标仪(BIO-TEK680).

表1 小鼠饲料配方质量比
1.2 方法
1.2.1 PAT氨基酸序列的获得以及过敏原蛋白数据库对比 PAT氨基酸序列的获得可在NCBI网站http://ncbi.nlm.nih.gov上查找到bar gene X17220,通过基因信息可得到其表达的磷丝菌素乙酰转移酶(phosphinothricin acetyl transferase,PAT)的全长氨基酸序列.然后,将PAT的氨基酸序列在三大数据库中进行比对:在SDAP上使用全序列比对、80 aa读码框滑行比对以及6 aa 框完全比对3 种算法进行预测;在Farrp使用全序列比对和80 aa 读码框滑行比对2 种算法预测;在NCBI 上进行全序列Blastp 分析.检索使用缺省设置值, 通过评估E-values、比对长度和一致性百分率来推断PAT是否具有致敏性或潜在致敏性[3].
1.2.2 PAT蛋白酶三级结构预测 将PAT蛋白的氨基酸序列输入SWISS-MODEL Workspace后,得到三维建模预测的蛋白结构,再通过PDB及PHYRE数据库分析系统可以找出与PAT蛋白酶相似的已知蛋白空间结构.通过分析已知的与PAT蛋白相似的蛋白酶的特性,推测出PAT酶是否有致敏性的可能.
1.2.3 动物饲养 60只小鼠随机等分为实验组和对照组,分别供应充足的饲料和水,保持鼠房的空气流通和稳定的温湿度(温度22~25 ℃,湿度50%~60%).亲代小鼠饲养90 d后,随机抽取每组小鼠各20只进行检测,其余用于繁殖F1代小鼠.F1代小鼠成年后即用于实验检测,F2代小鼠做同样处理.
1.2.3.1肠粘液sIgA的检测 解剖小鼠,打开腹腔,取其盲肠回部以上7 cm长小肠段,拭去血污,纵向平铺于滤纸上,手术剪挑开肠段,刮取小肠所有内容物与粘液于EP管中,加入1 mL PBS混匀.置于4 ℃冰箱保存.同样的方法处理其他小鼠.样本于4 ℃、离心机1 500 g离心20 min,取其上清.置于4 ℃冰箱保存待用.按照ELISA说明书步骤操作,并于450 nm处读出各孔吸光度(A值).
1.2.3.2 血清DAO和IgE的检测 小鼠摘除眼球放血,置于EP管中.静置1 h后,以4 ℃、1 000 g离心20 min.取上层血清,置冰箱中4 ℃温度下保存备用.按照ELISA说明书步骤操作,并于450 nm处读出各孔吸光度(A值).
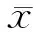
2 结果与分析
2.1 蛋白质生物信息学对PAT的比对和分析
将膦丝菌素乙酰转移酶(PAT)全序列183 aa找出后与三大蛋白数据库的已知过敏原进行比对后发现,PAT与已知致敏原无任何同源性(图1~5).
图1 膦丝菌素乙酰转移酶(PAT)全长氨基酸序列

图2 PAT蛋白序列在SDAP数据库比对搜索结果

图3 PAT蛋白序列与SDAP数据库过敏原的部分比对

图4 PAT蛋白序列与Farrp数据库过敏原全序列的部分比对

图5 PAT蛋白序列与Farrp数据库80 aa滑行比对结果
由以上各图可以看出,三大数据库的比对均显示PAT没有与已知过敏源相似的序列,这说明PAT酶的致敏可能性很低.
2.2 PAT蛋白酶三维结构预测及分析
将PAT蛋白酶的一级结构放入SWISS-MODEL三维结构建模系统进行解析,构建其可能的蛋白质结构;通过PDB及PHYRE数据库软件找出与之相似三级结构的蛋白质,一共发现有13种已知的或预测的蛋白质结构与其相似,均为N-酰基转移酶家族(N-acetyl Transferase Super Family,NAT SF)成员,PAT酶与常见的乙酰转移酶具有相似的三维构象特征(图6~7).它们共同的保守区域为乙酰辅酶A(acCoA)的化学绑定位点.
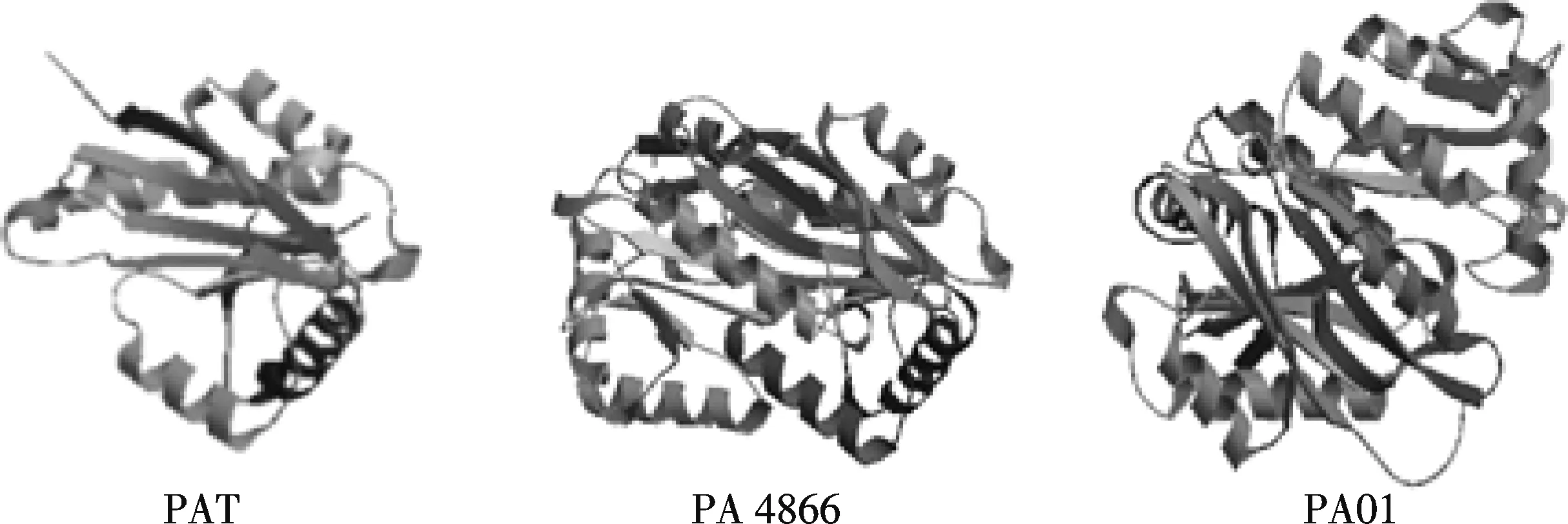
图6 PAT酶的三级结构预测、铜绿假单胞菌乙酰转移酶PA4866结构、铜绿假单胞杆菌乙酰转移酶PA01的预测结构

图7 PHYRE系统分析所得三维结构相似性蛋白图部分
2.3 sIgA、DAO和IgE含量检测
随机抽取各代小鼠每组20只,ELISA检测sIgA、DAO和IgE含量,用酶标仪于450 nm处读出各样本的吸光度OD值,即A值.SPSS17.0统计结果见表2.
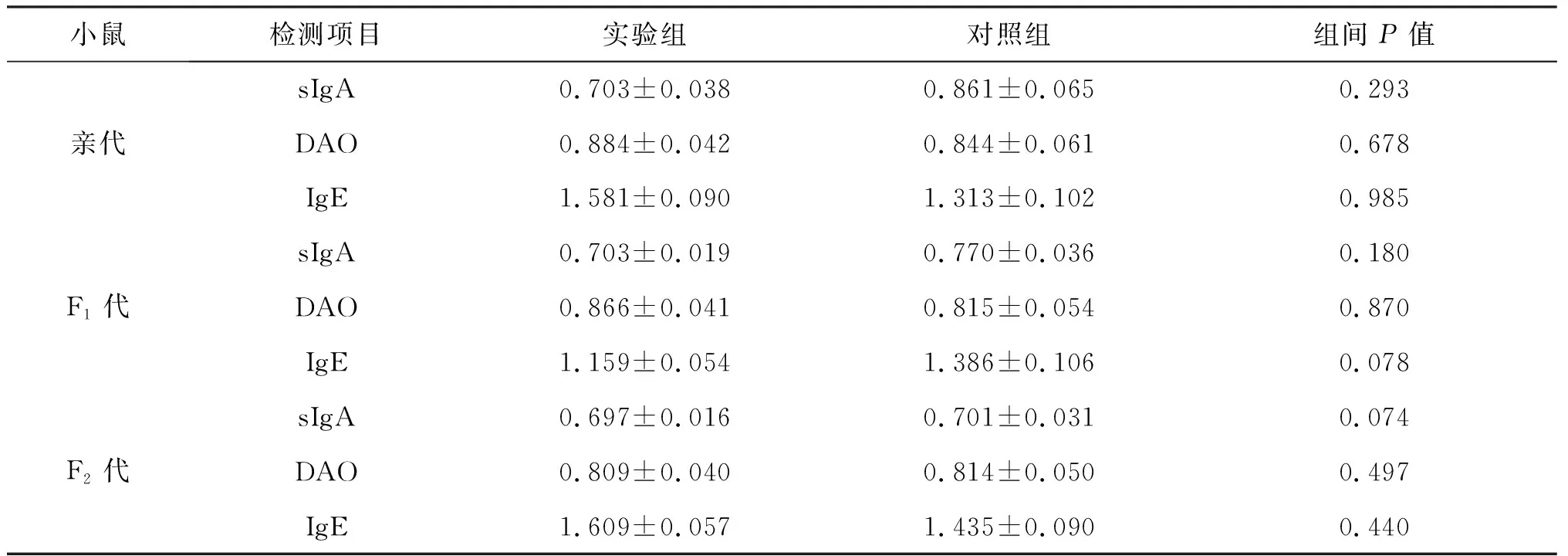
表2 sIgA、DAO和IgE吸光度(A值)统计分析(mean±SD)
由表2可以看出,亲代小鼠实验组肠道黏液sIgA的吸光度值(A值)略低于对照组,但描述性统计中的箱图显示其水平基本在对照组的范围之内,二者之间没有显著性的差异(0.703±0.038 vs 0.861±0.065,P=0.293>0.05).血清DAO的A值波动范围略大于对照组,中数略高,但是并没有显著性差异(0.884±0.042 vs 0.844±0.061,P=0.678>0.05).而血清IgE的组间比较时实验组出现了一个极大值,分析时用排除个案的方法排除此个案,再进行组间独立样本t检验,并没有显著性的差异(1.581±0.090 vs 1.313±0.102,P=0.985>0.05).推测这个极大值的出现可能系实验者的操作误差所造成,也有可能系小鼠自身产生了过敏反应,但考虑到过敏原的多重性以及IgE的特异性,IgE的增高还有可能是由于其他感染、炎症的发生,如鼠间的打斗撕咬导致伤口的感染发炎等.
F1代与F2代小鼠实验组与对照组相比, 肠道黏液中sIgA的含量、血清DAO含量、血清IgE含量与对照组也无显著性差异(P=0.180,0.178,0.078 >0.05;P=0.074,0.497,0.440>0.05).由此可见,食用转Bar基因稻谷并没有对小鼠的遗传产生影响,亲代实验组的后代并没有因为长期食用转基因稻谷而出现机体过敏增加.在90 d的观察期内,每一代小鼠都没有明显的过敏症状出现,如呕吐、腹泻、过敏性休克等.
3 小结与讨论
Bar基因(bialaphos resistance gene)来源于吸水链霉菌(streptamyces hygroscopicus),属于NAT Super Family中的一员.其作用机理为编码的PAT蛋白能使除草剂中草丁膦的自由氨基乙酰化而失去活性,从而呈现对除草剂的耐受性.吸水链霉菌在自然界中广泛存在,属于生物圈的一部分,链霉菌属中几乎没有任何菌种与人、动物、植物的病原体有关.吸水链霉菌与链霉菌属的许多菌种相似,表达蛋白PAT具有专一酶活性,可以认为这些菌种至少含有Bar基因的同源物,而目前尚无这些同源物对人和动物是毒素或过敏原的报道[11].
分析外源基因编码的氨基酸序列是判定食物是否致敏的最快捷方法,通常情况下致敏原具有结构和功能上的保守性,因此通过比对序列的同源性能够很好地对外源蛋白做出理论预测.本研究选用目前国际上过敏原相关信息比较丰富、功能强大并且可以互相补充的SDAP、Farrp和NCBI 三大数据库,进行PAT蛋白的氨基酸序列(图1)与已知过敏原相似性的比较,采用全序列比对、80 aa读码框滑行比对以及6 aa 框完全比对3种方法检测(图2~5).
目前蛋白序列相似性分析大多仅局限于比较氨基酸一级序列,由于除氨基酸顺序决定簇外还存在构象决定簇致敏,而序列相似性分析并不包括构象决定簇分析,因此,蛋白质一级结构的比较有一定局限性,对于非连续抗原决定簇的比较并不适用,而蛋白质的空间立体结构是保守的,所以比较其三维结构更为有效.本实验对PAT酶的三维构象进行了预测,并找出了与其氨基酸序列相似的蛋白的三维结构,均系NAT Super Family成员.这些乙酰转移酶家族的共同点是具有乙酰辅酶A(acCoA)的化学绑定位点这一保守区域(图6~7).
分泌型免疫球蛋白 A ( secretory Immunoglobulin A,sIgA )是黏膜免疫系统的主要体液防御因子, 许多食物过敏个体都会出现肠道黏膜损伤、肠壁通透性增加的现象.以sIgA作为主要防御因子的黏膜免疫系统含有特异的淋巴组织,它和内环境中的抗原接触并吞噬抗原,诱导T淋巴细胞、B淋巴细胞反应,B淋巴细胞分化的浆细胞分泌sIgA,起黏膜免疫保护作用[12-13].如若肠道粘液中sIgA的含量超出正常水平,则说明肠道免疫处于亢进状态,食物存在致敏性.二胺氧化酶(Diamine Oxidase,DAO)是一种具有高度活性的含有脱氨的腐胺和组胺的细胞内酶,是组胺等多胺物质的分解代谢酶,95%以上存在于哺乳动物小肠的黏膜或纤毛上皮细胞中,其活性与绒毛高度及肠黏膜细胞的核酸和蛋白合成密切相关.小肠黏膜屏障功能衰竭时,肠黏膜细胞脱落入肠腔,DAO进入肠细胞间隙淋巴管和血流,使血DAO升高.因此,血DAO活性可反映肠道损伤和修复情况.IgE 介导的免疫应答是食品过敏的主要效应,过敏原与IgE结合是过敏原发挥生物活性的中心环节[14],可以通过对IgE水平的检测来检验是否机体产生过敏反应.
本实验的sIgA、DAO检测结果无显著性差异,由此可见转Bar基因水稻并没有对小鼠肠道黏膜及免疫屏障造成损害,这可能是由于PAT蛋白对胃肠液无耐受性,在生物体的消化液中会快速降解掉.IgE的检测也未出现显著性差异,食用转Bar基因稻谷3代的小鼠都没有引起IgE介导的过敏反应,一方面可以说明转Bar基因的水稻无致敏性反应,但另一方面也可能跟Bar基因在水稻中的表达含量较低有关,虽然受试小鼠长期喂养且繁殖3代检测,但微量的PAT蛋白也不足以致敏.
参考文献:
[1] 张 婷. 关注转基因食物的潜在过敏性[J]. 免疫学杂志,2004, 20: 124-126.
[2] 林海霞, 常 艳, 梅其炳. 转基因作物食用潜在致敏性的安全评价[J]. 国外医学卫生学分册,2004, 34 (3):187-191.
[3] FAO/ WHO. Evaluation of allergenicity of genetically modifiedfoods[R]. Report of a Joint FAO/WHO Expert Consultation onAllergenicity of Foods Derived from Biotechnology. Food and Agriculture Organization of the United Nations (FAO), Rome, Italy 2001: 27-28.
[4] Codex Alimentarius Commission. Joint FAO/WHO Food Standard Programme, Codex Alimentarius Commission, Twenty-FifthSession [S]. Appendix III, Guideline for the Conduct of Food Safety Assessment of Foods Derived from Recombinant-DNA Plants and Appendix IV, Annex on the Assessment of Possible Allergenicity. Rome, Italy, 2003: 47-60.
[5] 赵杰宏, 韩 浩, 赵德刚. 转基因作物标记蛋白潜在致敏性的生物信息学预测 [J]. 中国烟草学报, 2010, 16(3): 76-79.
[6] 李英华, 朴建华, 卓 勤, 等. 转Xa21 基因大米亚慢性毒性实验[J]. 卫生研究, 2004, 33(5): 575-578.
[7] 陈小萍, 卓 勤, 朴建华, 等. 转基因大米的免疫毒理学评价[J]. 卫生研究, 2004, 33(1): 77-80.
[8] 周联高. 高赖氨酸转基因稻谷对肉仔鸡饲用安全性的研究[D]. 扬州:扬州大学, 2009.
[9] 黄 煌, 郑鹏远, 罗 予, 等. 食物过敏对小鼠肠道屏障功能及CD4+CD25+调节性T细胞的影响[J]. 世界华人消化杂志, 2008,16(17): 1932-1937.
[10] REEVES P G, ROSSOW K L, LINDLAUF J. Development and testing of the AIN-93 purified diets for rodents: results on growth, kidney calcification and bone mineralization in rats and mice[J]. J Nutrition,1993,123(11):1923-1931.
[11] HEROUET C, ESDAILE D J, Mallyon B A,etal. Safety evaluation of the phosphinothricin acetyltransferase proteins encoded by the pat and bar sequences that confer tolerance to glufosinate- ammonium herbicide in transgenic plants[J]. Regul Toxicol Pharmacol, 2005,41(2): 134-149.
[12] 王 魁. 分泌型 IgA及其在黏膜局部免疫抗感染中的研究现状[J]. 中国兽医科技, 2001, 20(8): 18-22.
[13] 杨定周, 周其全, 李素芝, 等. 高原缺氧致大鼠肠黏膜屏障功能损伤及谷氨酰胺的保护作用观察[J]. 解放军医学杂志, 2011, 36(3): 301-306.
[14] KLETER G A, PEIJNENBURG A A. Screening of transgenic proteins expressed in transgenic food crops for the presence of short amino acid sequences identical to potential, IgE-binding linear epitopes of allergens[J]. BMC Struct Biol, 2002, 2(1):2-8.
