食品中热损伤沙门氏菌选择性增菌及实时荧光聚合酶链式反应检测
2012-10-25滕要辉史贤明艾志录
索 标,滕要辉,史贤明,艾志录
(1.河南农业大学食品科学技术学院,河南 郑州 450002;2.上海交通大学农业与生物学院,上海 200240)
食品中热损伤沙门氏菌选择性增菌及实时荧光聚合酶链式反应检测
索 标1,2,滕要辉1,史贤明2,艾志录1
(1.河南农业大学食品科学技术学院,河南 郑州 450002;2.上海交通大学农业与生物学院,上海 200240)
为了能够高效检测食品中的亚致死损伤沙门氏菌,首先采用热激胁迫的方法得到亚致死损伤沙门氏菌细胞,进而基于选择性增菌培养液SEL建立一种实时荧光聚合酶链式反应检测技术。结果表明:在SEL中经过20h增菌培养后,无论是否经过修复培养,1~2CFU/5mL SEL的亚致死损伤细胞都能得到完全修复并增菌至109CFU/mL水平;依此建立的实时荧光聚合酶链式反应检测技术的纯菌扩增效率为95.41%,检测限为4CFU/反应体系,在人工污染碎食品样品中的检测限为3CFU/10g碎牛肉,而且与传统培养检测方法的结果相吻合。本方法在24h内即可完成食品中热损伤沙门氏菌的修复、选择性增菌以及实时荧光聚合酶链式反应检测,可应用于食品中沙门氏菌污染状况调查及高效检测。
热损伤;沙门氏菌;选择性增菌;实时荧光聚合酶链式反应;检测
沙门氏菌(Salmonella)是一种广泛分布于自然界的危害极大的肠道致病菌,在我国,40%的细菌性食物中毒病例是由沙门氏菌引起的[1],因此必须采取高效的检测手段以保障食品的安全。实时荧光聚合酶链式反应(polymerase chain reaction,PCR)方法是检测微生物的有效分子手段之一[2],现在已经建立一些实时荧光PCR检测技术以用于食品中沙门氏菌的检测[3]。
食源性致病菌经过热激等物理控制方法处理后会出现3种生理上状态不同的细胞——未受损伤的细胞、亚致死损伤细胞和死亡细胞[4]。由于食品中亚致死损伤细胞存在的可能性,新型分子检测技术的开发需要能够同时检出活性细胞和损伤细胞[5]。此外,绝大多数食品样品中的致病菌都是以很低的水平存在(每25g食品一般只有1~2个细胞),食品样品中也存在大量的背景微生物菌群,因此,在检测之前仍需要对食品中的致病菌进行选择性增菌培养[6]。然而,目前常用的作为选择性培养基的成分对损伤致病菌的修复和生长会产生抑制作用[7],这也是目前食源性致病菌检测之前,大都需要经过非选择性修复培养后再经过选择性增菌这两步的主要原因[8]。SEL是Kim等[9]开发出来的用于沙门氏菌、大肠杆菌O157∶H7以及单核细胞增生李斯特菌的选择性增菌培养液,本人前期也采用SEL建立了一种多重实时荧光PCR检测平台[10],但目前对该培养基修复并增菌培养损伤致病菌的能力尚不清楚。
本研究以鼠伤寒沙门氏菌为供试菌株,采用热激的方法得到亚致死损伤细胞,进而研究其在SEL中的修复能力,建立实时荧光PCR检测技术,并在人工污染的牛肉样品中进行检测试验,从而建立一种新型的基于选择性培养基SEL的损伤沙门氏菌的修复、选择性增菌和实时荧光PCR检测平台。
1 材料与方法
1.1 材料、试剂与仪器
鼠伤寒沙门氏菌(Salmonella entericaTyphimurium ATCC14028) 美国标准菌种收藏所。
木糖赖氨酸去氧胆酸盐琼脂(XLD)、营养肉汤(NB)、营养肉汤琼脂(NA)、营养肉汤琼脂(NASA,添加4% NaCl)、缓冲李氏菌增菌肉汤(BLEB) 美国BD公司。
DNeasy组织总DNA提取试剂盒 美国Qiagen公司;抗生素Acriflavine、Nalidixic acid sodium salt、Phosphomycin disodium salt以及 Cycloheximide C 美国Sigma公司;荧光定量PCR试剂盒 美国应用生物系统公司;引物和实时荧光PCR探针由美国应用生物系统公司合成。
恒温混合器、移液器 德国Eppendorf公司;ABI 7500型实时荧光PCR仪 美国应用生物系统公司;NanoDrop ND-100分光光度计 美国NanoDrop科技公司。
1.2 方法
1.2.1 热损伤沙门氏菌的制备
将鼠伤寒沙门氏菌ATCC14028于37℃培养3.5h至指数后期,取0.5mL于55℃热激处理6min后,热激亚致死损伤细胞立即用于下步操作。
1.2.2 亚致死沙门氏菌在选择性增菌培养液SEL中的修复
将热激后的亚致死沙门氏菌菌液用0.9%生理盐水进行梯度稀释,分别在NA、XLD和NASA上测定活性细胞的数目。取50μL梯度稀释后的亚致死菌液分别加入到有5mL BLEB培养基的20mL PA瓶中,以无菌0.9%生理盐水作为空白对照,37℃、150r/min振荡修复培养0、1h和2h后,根据选择性增菌液SEL的配方[9],加入各种抗生素,以得到终质量浓度分别为0.01g/L的Acriflavine、0.05g/L的Cycloheximide、0.05g/L的Fosfomycin以及0.002g/L的Nalidixic acid。同等条件下再培养6h和20h后,从每个处理中各取1mL用于活细胞计数。每样重复3次,取平均值。
1.2.3 亚致死损伤沙门氏菌细胞数目的测定
参照Aljarallah等的方法进行[11],将热激或SEL培养后的菌液经10倍系列稀释后,取0.1mL涂布于NA、XLD和NASA平板上,细胞外膜受损的亚致死细胞无法在XLD培养上生长,而细胞质膜受损的亚致死细胞无法在NASA培养基上生长,而亚致死损伤细胞在NA培养基上可以正常生长[11]。涂布后的NA和XLD平板在37℃培养24h,NASA平板培养48h,预实验也证明更长的培养时间对存活细胞的计数结果没有任何影响。培养结束后,计数各培养基上的菌落形成单位(CFU),根据各试验结果,计算平均值和标准偏差。
1.2.4 基因组DNA的提取
1mL的细菌培养液用8000×g离心10min收集细胞后,用QIAGEN DNeasy Tissue Kit试剂盒法提取基因组DNA。提取后的基因组DNA用NanoDrop ND-100分光光度计确定所提取基因组DNA溶液的浓度与纯度。
1.2.5 实时荧光PCR
参照前期建立的实时荧光PCR方法[10],所用invA基因引物的特异性已经得到证明。上游引物序列为:GTTGAGGATGTTATTCGCAAAGG;下游引物序列为:GGAGGCTTCCGG GTCAAG;TaqMan探针序列为:CCGTCAGACCTCTGGCAGTACCTTCCTC,探针的3′端标记有FAM荧光染料。每20μL实时荧光PCR反应体系中包含1×缓冲液、200nmol/L引物和探针、以及2μL的基因组DNA。混匀后,加入96孔板中,再加入2μL待检测的模板基因组DNA,置实时荧光PCR仪上反应。反应程序为:95℃热启动10min,随后是40个循环的95℃变性15s以及60℃复性和延伸1min。
1.2.6 亚致死沙门氏菌修复与实时荧光检测技术在人工污染食品样品中的应用
将0.1mL梯度稀释后的亚致死鼠伤寒沙门氏菌ATCC 14028分别人工污染于10g碎牛肉样品中,供试碎牛肉购自附近超市,并预先用传统培养的方法(《USDA/FSIS Microbiology Laboratory Guidebook》)验证样品中不含有沙门氏菌的污染。在SEL选择性增菌培养液中增菌培养20h后,1mL增菌液用于基因组DNA的提取和实时荧光PCR检测,同时,样品中修复并增菌后沙门氏菌的数目也用XLD选择性平板计数法测定,以作为该实时荧光PCR检测方法的对照。人工污染试验重复进行3次,每次包含两个平行处理。
2 结果与分析
2.1 热损伤沙门氏菌在选择性培养基SEL中的修复
本试验用选择性增菌培养基SEL回复培养55℃热激亚致死损伤的沙门氏菌,结果如表1所示,热激后的起始各浓度沙门氏菌细胞在XLD上的活细胞数目较在NA上少,而NASA平板上均不可生长,说明部分细胞的外膜受到了损伤,而几乎全部活性细胞的质膜均受到了不同程度的损伤,为亚致死损伤细胞[11]。在SEL中增菌培养后的结果表明,如果接种量大于1.80×102CFU/5mL SEL时,在亚致死损伤细胞经过2h的非选择性修复培养后,再在加入选择性抗生素的SEL中经过6h的增菌培养,在XLD和NASA培养基上能检出与在NA培养基上大致相等的菌落数目,如果只经过1h的回复培养,则1.80×102CFU/5mL SEL接种量的PA瓶中的菌液在NASA上无活性细胞检出。而没有经过回复培养的细菌,经过6h的增菌培养后,1.80×102CFU/5mL SEL或更低接种量的菌液在3种培养基上均无活性细胞检出,更高接种量的菌液经过6h的SEL培养后,在3种培养基上的活性细胞数目基本一致,但与初始接种量相比,细胞数目并没有得到明显的增加。在SEL中经过20h的增菌培养后,无论是否经过一定时间的修复培养,各接种量菌液中的细胞都能得到完全修复并增菌到109CFU/mL。
2.2 实时荧光PCR检测灵敏度
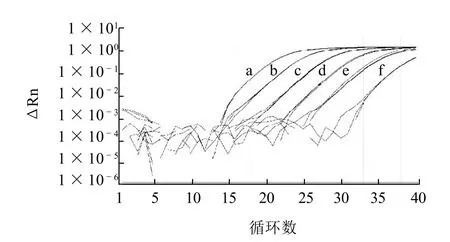
图1 沙门氏菌实时荧光PCR检测灵敏度Fig.1 Detection sensitivity of Salmonella by Real time PCR
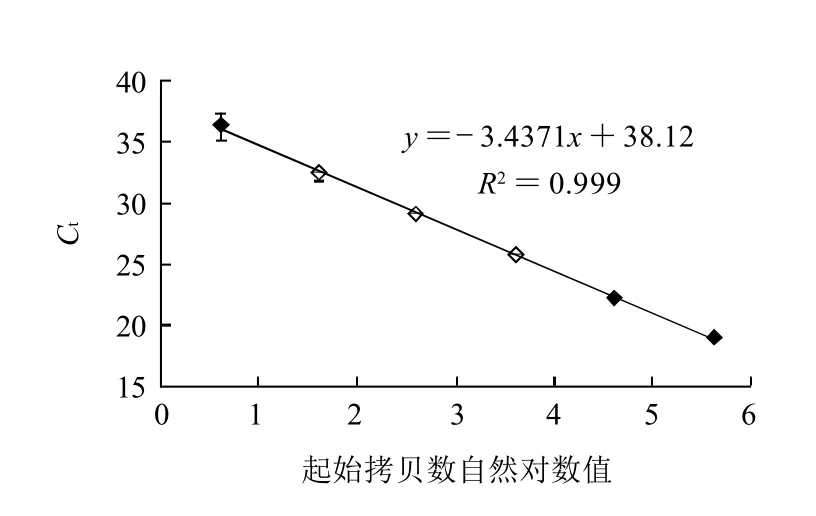
图2 沙门氏菌实时荧光PCR检测效率Fig.2 Detection efficiency of Salmonella by Real time PCR
为了测定实时荧光PCR检测沙门氏菌的扩增效率,采用鼠伤寒沙门氏菌ATCC14028作为供试菌株,根据不同浓度的基因组DNA的扩增结果计算出标出曲线,再根据标准曲线的斜率k(-3.4371),由公式E=(10-1/k)-1计算出扩增效率(E)。如图1、2所示,各质量浓度基因组DNA扩增曲线之间距离均匀,扩增效率为95.41%,最低为4个拷贝数的沙门氏菌基因组DNA可被检测出。
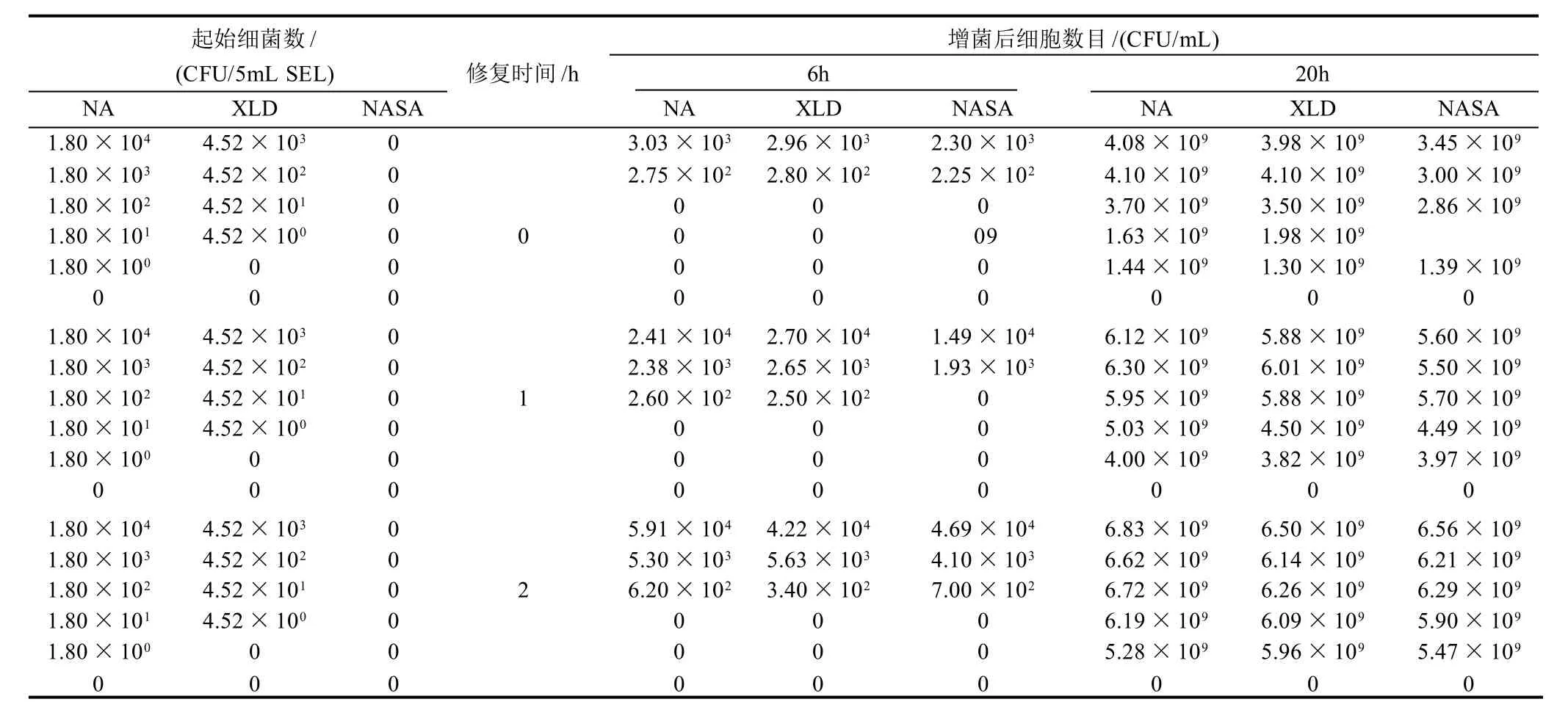
表1 热损伤沙门氏菌在SEL培养液中的修复和增菌培养结果Table 1 Results of recovery and enrichment of thermal injured Salmonella in SEL broth
2.3 实时荧光PCR检测方法在人工污染碎牛肉样品中的应用
一系列10倍梯度稀释的热激亚致死鼠伤寒沙门氏菌ATCC 14028接种于10g碎牛肉中,经过在SEL中20h的增菌培养后,增菌液用于实时荧光PCR检测,同时用选择性XLD平板对各致病菌进行计数作为对照。结果如表2所示,经过20h的增菌培养后,用这两种检测方法均能从10g碎牛肉样品中检测出最低起始数目为3CFU的沙门氏菌,然而, 实时荧光PCR定量检测结果得到的细胞数目略高于平板菌落计数结果。

表2 实时荧光PCR结合SEL选择性增菌法从人工污染碎牛肉样品中检测沙门氏菌Table 2 Real-time PCR combined with SEL broth for detection of Salmonella in spiked ground beef
3 讨 论
细菌受到热激处理后会出现亚致死损伤甚至死亡,亚致死致病菌的检测也是食品安全研究检测的热点和难点之一[5]。食品中亚致死损伤细胞的存在为食源性致病菌安全检测提出了新的要求,必须引起人们的足够重视,因为在食源性致病菌检测过程中,亚致死损伤细胞的存在有可能会高估致病菌的数目[12],也有可能会造成假阴性检测结果,特别是当用选择性培养基检测致病菌的时候[13],在后面一种情况下,必须包括一个亚致死损伤细胞的修复培养步骤。
目前用于亚致死损伤致病菌修复培养的培养基大多采用不含选择性成分的营养培养基,如BPW、UPB以及TSB等[5,14]。曾庆梅等[15]研究了不同介质、温度和时间对热损伤肠炎沙门氏菌的修复的影响,得出最佳修复条件为营养肉汤37℃修复2h。然而,这些培养基均无法满足检测目标致病菌选择性增菌的需要。由于食品样品中大量背景菌群的存在,选择性增菌培养也是避免出现假性检测结果的重要步骤[16]。本实验首次评价了选择性增菌培养液SEL修复以及增菌培养热致亚致死损伤鼠伤寒沙门氏菌的能力,结果表明了该培养液除了能够选择性增菌培养沙门氏菌之外[9],也能够很好的修复鼠伤寒沙门氏菌外膜和质膜的损伤关键部位,但与前人报道的非选择性培养液相比[17],添加了抗生素的该选择性培养基也明显延缓了细胞生长速度,既使经过1~2h的修复培养,6h的增菌培养也无法将1.80×101CFU/5mL SEL或更低的细胞增菌至可检出水平,因此,20h的增菌培养对于后续的分子生物学检测,仍是亚致死损伤沙门氏菌修复和增菌至可检出水平所必须的。在该增菌培养条件下,即使不经过非选择性修复步骤,1~2CFU/5mL SEL的热激亚致死沙门氏菌也能够增菌至可检出水平。
在人工污染的碎牛肉样品中,结合SEL选择性增菌培养液,经过20h的培养,本实时荧光PCR检测方法能在大量背景菌存在的条件下,从10g碎牛肉样品中修复、增菌并检测出2~3CFU的沙门氏菌,这也是目前实时荧光PCR等分子生物学方法所能达到的最低检测限[16]。
本研究采用了活性平板计数的方法对增菌后的人工污染碎牛肉样品中的致病菌进行了计数,并与实时荧光PCR的定量结果进行了比较,结果表明在所得到的分别用CFU/mL与基因组DNA拷贝/mL表示的定量结果之间有一定的差异,这是因为活细胞技术的方法仅仅可以测定能在选择性平板上生长的具有活性的细胞,而PCR的方法定量的结果则包括了所有可培养以及不可培养的细胞,因而在食品样品中检测出的基因拷贝数结果大多高于活细胞数目。
至于该实时荧光PCR的检测速率,从样品增菌到检测结果的分析,整个检测过程可在24h内完成。与传统的以培养为基础的方法所需要的5~7d才能得到结果相比,本研究所开发的实时荧光PCR方法显著提高了食品中沙门氏菌检测的效率。
总之,本研究基于选择性增菌培养液SEL,开发出了一个快速、灵敏的实时荧光PCR方法以从食品样品中选择性修复、增菌并检测出热损伤沙门氏菌。经过对其检测灵敏度和有效性的综合评价,本检测方法可以完成大量食品样品中包括亚致死细胞在内的沙门氏菌高效快速检测。
[1] 童哲, 程苏云, 梅玲玲. 浙江省272份食品沙门氏菌检测结果[J]. 浙江预防医学, 2003, 15(4)∶ 33-34.
[2] ESPY M J, UHL J R, SLOAN L M, et al. Real-time PCR in clinical microbiology∶ applications for routine laboratory testing[J]. Clin Microbiol Rev, 2006, 19 (1)∶ 165-256.
[3] 麻丽丹, 王殿夫, 巴中华, 等.TaqMan MGB PCR定量检测水产品中沙门氏菌的方法建立[J]. 食品科学, 2009, 30(20)∶ 303-307.
[4] WUYTACK E Y, PHUONG L D, AERTSEN A, et al. Comparison of sublethal injury induced inSalmonellaenterica serovarTyphimuriumby heat and by different nonthermal treatments[J]. J Food Prot, 2003, 66(1)∶31-37.
[5] WU V C. A review of microbial injury and recovery methods in food[J].Food Microbiol, 2008, 25(6)∶ 735-744.
[6] MANDAL P K, BISWAS A K, CHOI K, et al. Methods for rapid detection of foodborne pathogens∶ an overview[J]. Am J Food Technol,2011, 6(2)∶ 87-102.
[7] JACOBSEN C N. The influence of commonly used selective agents on the growth ofListeria monocytogenes[J]. Int J Food Microbiol, 1999,50(3)∶ 221-226.
[8] NUGEN S R, BAEUMNER A J. Trends and opportunities in food pathogen detection[J]. Anal Bioanal Chem, 2008, 391(2)∶ 451-454.
[9] KIM H, BHUNIA A K. SEL, a selective enrichment broth for simultaneous growth ofSalmonella enterica,Escherichia coliO157∶H7, andListeria monocytogenes[J]. Appl Environ Microbiol, 2008, 74(15)∶4853-4866.
[10] SUO Biao, HE Yiping, TU S I, et al. A multiplex real-time polymerase chain reaction for simultaneous detection ofSalmonellaspp.,Escheri-chia coliO157, andListeria monocytogenesin meat products[J].Foodborne Pathog Dis, 2010, 7(6)∶ 619-628.
[11] ALJARALLAH K M, ADAMS M R. Mechanisms of heat inactivation inSalmonellaserotypeTyphimuriumas affected by low water activity at different temperatures[J]. J Appl Microbiol, 2007, 102(1)∶ 153-160.
[12] RITZ M, PILET M F, JUGIAU F, et al. Inactivation ofSalmonella TyphimuriumandListeria monocytogenesusing high-pressure treatments∶destruction or sublethal stress?[J]. Lett Appl Microbiol, 2006, 42(4)∶357-362.
[13] HOEFEL D, GROOBY W L, MONIS P T, et al. Enumeration of waterborne bacteria using viability assays and flow cytometry∶ a comparison to culture-based techniques[J]. J Microbiol Met, 2003, 55(3)∶ 585-597.
[14] JIANG J, LARKIN C, STEELE M, et al. Evaluation of universal preenrichment broth for the recovery of foodborne pathogens from milk and cheese[J]. J Dairy Sci, 1998, 81(11)∶ 2798-2803.
[15] 曾庆梅, 张冬冬, 韩抒, 等. 热损伤肠炎沙门氏菌的修复方法研究[J].食品科学, 2009, 30(23)∶ 233-236.
[16] 索标, 汪月霞, 艾志录. 食源性致病菌多重分子生物学检测技术研究进展[J]. 微生物学杂志, 2010, 30(6)∶ 71-75.
[17] CHEN Jing, ZHANG Lida, PAOLI G C, et al. A real-time PCR method for the detection ofSalmonella entericafrom food using a target sequence identified by comparative genomic analysis[J]. Int J Food Microbiol,2010, 137(2/3)∶ 168-174.
Selective Enrichment and Real time PCR Detection of Thermal Injured Salmonella in Food
SUO Biao1,2,TENG Yao-hui1,SHI Xian-ming2,AI Zhi-lu1
(1. College of Food Science and Technology, Henan Agricultural University, Zhengzhou 450002, China;
2. School of Agriculture and Biology, Shanghai Jiao Tong University, Shanghai 200240, China)
A selective enrichment broth, SEL based real time PCR assay was established after treating a sublethal injuredSalmonellacells by heat-shock. The results showed that 1-2 CFU of injuredSalmonellacells in 5 mL SEL could be fully recovered and enriched to the level of 109CFU/mL after a 20 h enrichment step even without the nonselective recovery process. The amplification efficiency of real time PCR was 95.41% and the detection limit was 4 CFU/reaction forSalmonellagenomic DNA.The detection limit in artificially contaminated food sample was 3 CFU/10 g ground beef, and showed a high coincidence to the detection result by traditional culture-based assay. The overall recovery, selective enrichment and detection procedure of thermal injuredSalmonella could be accomplished in 24 h, which showed a high potential of application in the effective detection ofSalmonellain food.
thermal injury;Salmonella;selective enrichment;real time polymerase chain reaction (PCR);detection
TS254.1
A
1002-6630(2012)10-0223-05
2011-04-17
河南农业大学(人才引进/博士启动)基金项目(30300168);河南省重点科技攻关计划项目(082102320006)
索标(1982—),男,副教授,博士,研究方向为食品微生物分子生态与食品安全。E-mail:suobiao@gmail.com
