双麦麦精对嗜热链球菌生长影响的研究
2012-10-25刘少伟陈如琪吕宝龙卢艳花
赵 爽,刘少伟,*,陈如琪,吕宝龙,卢艳花,费 楠
(1.华东理工大学,上海200237;2.上海英联食品饮料有限公司,上海 200245)
双麦麦精对嗜热链球菌生长影响的研究
赵 爽1,刘少伟1,*,陈如琪1,吕宝龙1,卢艳花1,费 楠2
(1.华东理工大学,上海200237;2.上海英联食品饮料有限公司,上海 200245)
主要研究双麦麦精对嗜热链球菌生长过程的影响。通过在营养成分缺省的培养基中添加双麦麦精,采用分光光度法测定菌液吸光值(OD600),根据OD值确定对应的嗜热链球菌的活菌数,从而分析双麦麦精对嗜热链球菌生长情况的影响。利用正交实验,通过直接计数法确定双麦麦精与其他营养成分的最佳比例。结果表明,双麦麦精对嗜热链球菌的生长有明显的促进作用,可为嗜热链球菌提供充足的碳源和生长因子。双麦麦精的添加量为16g/L时效果最佳。分析比较吸光值法和直接计数法测得的生长曲线,结果表明两种方法具有良好的相似性,其中在对数期有强相关性。
嗜热链球菌,生长曲线,双麦麦精,缺省培养基,菌落计数
近十年来,随着健康意识的增强和人民生活水平的提高,我国发酵乳制品工业迅猛发展[1]。发酵乳制品中的乳酸菌具有良好的保健作用,产品营养价值较高,并能改善肠道正常微生物区系的稳定性[2]。乳酸菌数量是评价该类产品营养价值与保健作用的重要依据[3-5]。嗜热链球菌作为一种对人体有明显食疗作用的有益菌,是生产酸奶的重要微生物菌群[6]。以大麦为原料制备的麦精和麦芽汁作为培养基原料已有广泛的应用[7-9]。双麦麦精是以大麦,其他麦类为原料通过酶解提取的一种天然营养物质。前期研究表明,在酸奶制作过程中添加双麦麦精,对酸奶中保加利亚杆菌、嗜热链球菌以及双歧杆菌的生长有明显的促进作用,并且在其储存阶段起到一定的保护作用[10]。在Domiati奶酪中添加麦精可以促进奶酪的香味,使奶酪有更好的蛋白水解和脂肪分解并具有更高的菌体含量[11]。麦精具有抗氧化性[12],其具有多种功能性成分,β-葡聚糖就是研究较多的一种益生元,此外还有低聚糖、生育三烯酚、γ-氨基丁酸等[13]。现已证明低聚糖对人体健康具有重要作用,并对提高乳酸菌活菌数具有促进作用[14-15],能增强乳酸菌的抗冷冻性和抗胆汁能力[16-17]。本文在前期工作的基础上,用含有双麦麦精的培养基和培养液对嗜热链球菌进行培养,观察双麦麦精对嗜热链球菌生长的影响,探讨双麦麦精作用于嗜热链球菌的意义。并分析分光光度法和直接计数法两种嗜热链球菌生长曲线测定方法的差异,分析其相关性,根据所得结果可对嗜热链球菌的培养过程进行指导。
1 材料与方法
1.1 材料与仪器
嗜热链球菌 YO-MIXTM499(Streptococcus thermophillus) 华东理工大学生物科学与工程学院食品系实验室提供;双麦麦精 上海英联食品饮料有限公司提供,双麦麦精主要成分为蛋白质7%,还原糖28%;M17培养基(g/L) 酵母膏2.5,蛋白胨10.0,牛肉膏5.0,乳糖5.0,抗坏血酸0.5,磷酸氢二钾5.0,硫酸镁0.5,甘油10.0,调pH至7.0~7.2。
超净工作台SW-CJ-2FDA 苏州安泰空气技术有限公司;电热恒温培养箱DHP-9052 上海一恒科学仪器有限公司;高压蒸汽灭菌器SYQ-DSX-280B
上海申安医疗器械厂;pH计FE20型 梅特勒-托利多公司;紫外可见分光光度计UV-2000 尤尼柯公司。
1.2 实验方法
1.2.1 菌种活化 保存的菌种活力不旺盛,处于维持生命的休眠状态,因而在使用前要进行活化。嗜热链球菌活化方法:将4℃冰箱中保存的嗜热链球菌菌种以质量分数3%的接种量接入灭菌牛奶中,按照嗜热链球菌的生长最适温度,放入42℃的培养箱中培养24h,待凝乳后取出,放在冰箱中保存,每7d活化一次,活化三代。
1.2.2 缺省培养基配制 以M17为基本培养基,根据双麦麦精为嗜热链球提供不同营养成分(碳源、氮源、生长因子)设计培养基,见表1。
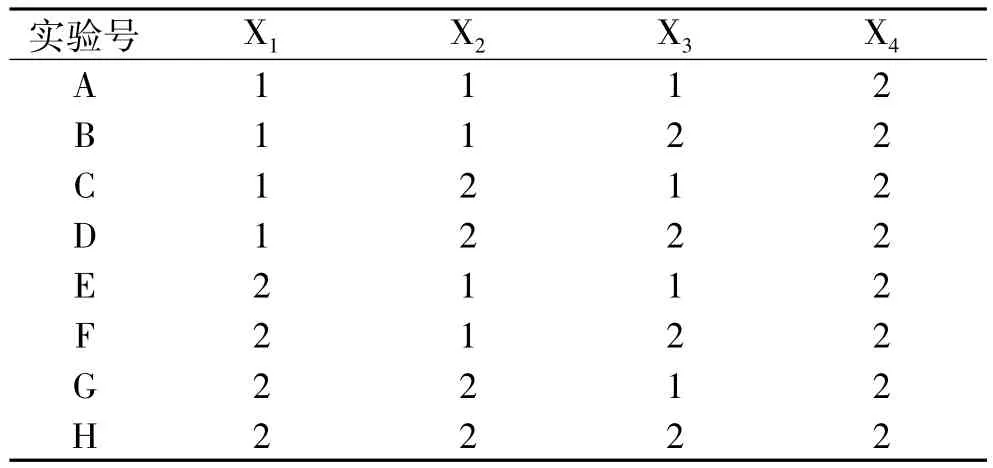
表1 培养基的配制Table 1 Preparation of medium
1.2.3 发酵培养 将活化过的菌种按质量分数3%接种量加入到M17培养基和缺省培养基中,放入42℃的培养箱中培养,进行OD600值和活菌数的测定。
1.2.4 菌液吸光值(OD600)的测定 摇床培养温度为42℃,摇床转速为180r/min,使用分光光度计,在600nm条件下通过比色法测定菌液的吸光值,从0h开始,每2h测定一次,测定12次。用未接种的培养基适当稀释后作空白。以培养时间为横坐标,相应的吸光值为纵坐标,3次平行实验,求平均值,绘制生长曲线。
1.2.5 活菌落计数方法 采用逐级稀释倒平板及涂布法两种。取菌液1mL,用灭菌蒸馏水依次稀释10、102、103、104、105倍,选取菌落数在30~300个之间进行计数,取菌液0.1mL分别滴于不同培养基上,42℃培养72h。72h后统计菌落数,相同培养基上的菌落数以平均值为准记数。
1.2.6 正交实验 为了确定双麦麦精与其他营养成分的最佳比例,在缺省成分实验的基础上,以由正交实验碳源浓度(A)、氮源浓度(B)、双麦麦精浓度(C)三个因素,每个因素选取三个水平,进行L9(34)正交实验。

表2 L9(34)正交实验因素水平表Table 2 L9(34)orthogonal experimental level table
1.3 分析方法
本实验所有数据使用SPSS软件进行数据分析,并用Excel进行制图。
2 结果与分析
2.1 嗜热链球菌在缺省培养基中的生长曲线比较
2.1.1 同时缺省碳源和氮源的生长情况 由图1可知,嗜热链球菌在缺省碳源和氮源的A、B培养基的前0~10h,长期处于生长的调整期,前8h与M17培养基生长情况类似,之后生长缓慢。与M17培养基的生长曲线相比,调整期延长了4h。培养终点24h,A、B培养基下的OD600值仅为M17的一半,说明双麦麦精不能为嗜热链球菌生长提供充足的碳源和氮源,嗜热链球菌不能仅靠双麦麦精进行大量增殖。A、B培养基的生长曲线差异不显著(p>0.05),双麦麦精在这种环境下,完全可以作为嗜热链球菌的生长因子,达到和原生长因子(抗坏血酸、硫酸镁、甘油、磷酸氢二钾)同样的效果。
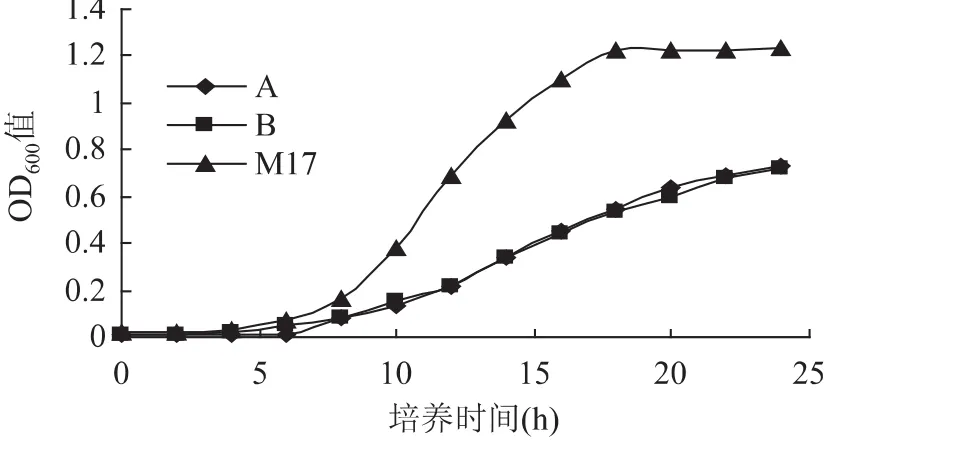
图1 缺省碳源和氮源的嗜热链球菌生长曲线Fig.1 Lack of source of carbon and nitrogen source of streptococcus thermophilus cultured growth curve
2.1.2 缺省碳源的生长情况 由图2可知,嗜热链球菌在缺省碳源的C、D培养基中的生长情况与M17中类似,在处于调整期的0~8h生长情况基本一致。从对数期开始,C、D的OD600值略高于M17的培养基,但C、D培养基的嗜热链球菌比M17的稍晚进入对数期。说明双麦麦精可以为嗜热链球菌提供充足的碳源,并且效果略优于M17培养基中的碳源成分(乳糖)。但在双麦麦精作为单一碳源的情况下,进入对数期较慢,可能由于嗜热链球菌分解乳糖的速度快于分解麦芽糖,因而乳糖更容易被嗜热链球菌吸收利用,而在对于双麦麦精中麦芽糖的吸收利用,需要更多的时间。此条件下,C、D生长曲线差异不显著(p>0.05),说明在此条件下,双麦麦精可以提供嗜热链球菌生长需要的生长因子。
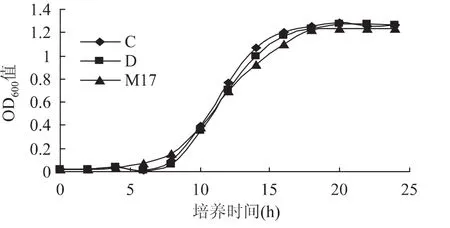
图2 缺省碳源的嗜热链球菌生长曲线Fig.2 Lack of carbon source streptococcus thermophilus cultured growth curve
2.1.3 缺省氮源的生长情况 由图3可知,嗜热链球菌在缺省氮源的E、F培养基中的处于调整期的时间较短,比M17中的嗜热链球菌提早2h进入对数期,也明显早于C、D培养基。在6~8h的对数期,双麦麦精可提供一定量的嗜热链球菌生长所需的氮源,但在8~16h,随着嗜热链球菌数量的增长,氮源的需求不能得到满足,相对不足,造成了对数期整体时间短,增长速率慢的结果。最终在稳定期的OD600值远小于M17培养基,说明双麦麦精在作为嗜热链球菌氮源的能力比较有限,如果要通过双麦麦精获得足够的氮源,还需要增加双麦麦精的添加量。在此条件下,双麦麦精仍可以提供嗜热链球菌生长需要的生长因子,最终限制嗜热链球菌生长的原因是氮源不足。
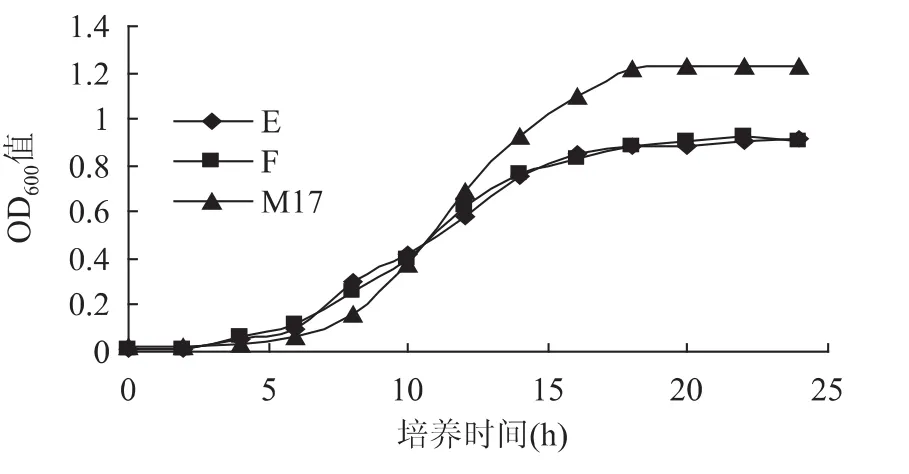
图3 缺省氮源的嗜热链球菌生长曲线Fig.3 Lack of nitrogen sourcestreptococcus thermophilus cultured growth curve
2.1.4 缺省生长因子的生长情况 由图4可知,嗜热链球菌在缺省生长因子的G培养基中可以快速增殖,在稳定期内的OD600值大于M17培养基,但略微少于无缺省培养基H。说明一方面,双麦麦精能为嗜热链球菌提供原M17培养基中没有的生长因子,对嗜热链球菌的生长产生了积极作用。另一方面双麦麦精也能为嗜热链球菌提供部分的碳源和氮源,保证了嗜热链球菌生长在营养充足的环境中。H培养基达到最高菌数的时间比M17培养基大约提前了4h,在稳定期的OD600值也是所有培养基中最高的。说明在提供双麦麦精使其所有营养条件充足的情况下,可以有效的缩短嗜热链球菌调整期和对数期的时间,使其提前到达稳定期。
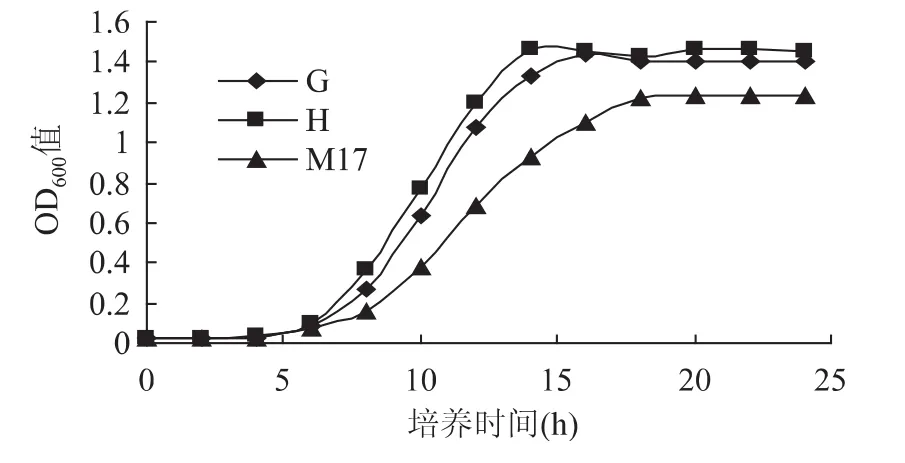
图4 缺省生长因子的嗜热链球菌生长曲线Fig.4 Lack of growth factor Streptococcus thermophilus cultured growth curve
2.2 正交实验
正交实验的结果(表3)表明:最终成分优化的结果为A2B3C3,即乳糖浓度5g/L,酵母膏浓度4g/L,蛋白胨浓度12g/L,牛肉膏浓度7g/L,双麦麦精浓度16g/L。在此条件下,测得嗜热链球菌活菌数为35.9 ×107cfu/mL。同时通过比较各项指标的极差R值,可以得到各因素的主次关系为B>A>C。

表3 培养基成分正交实验结果Table 3 Orthogonal experiment results
作为对照的 M17培养基的活菌数为3.03× 108cfu/mL。如果培养基双麦麦精浓度较低,则培养基营养相对不足,嗜热链球菌的生长就会受到一定的限制。如果培养基双麦麦精浓度过高,不仅不会促进细胞增长,反而会抑制菌体增殖,因为浓度过高,造成整个培养基的浓度过大,产生细胞渗透现象,造成嗜热链球菌失水皱缩,导致生长受到影响。不同双麦麦精添加量的菌落均为乳白色,光滑,边缘整齐,直径约2mm,菌落较厚,无明显差异。
2.3 生长曲线比较
2.3.1 分光光度法和直接计数法整体比较 嗜热链球菌在适宜生长条件下菌体细胞生长出现4个阶段:调整期、对数期、稳定期、衰亡期。生长曲线反映了嗜热链球菌在培养过程中的繁殖规律,同一嗜热链球菌在不同培养条件下的生长曲线不同,不同测定方法测得的生长曲线也不同。目前,主要采用分光光度法和直接计数法测定生长曲线。前者所测为包含已死亡菌体和因菌体自溶产生的吸光值,并且据朗伯—比尔定律,各类低浓度溶液吸光值定量测定的依据,浓度过高或过低均影响测量结果的准确性。后者所测为实际观察并计数得到的菌体细胞数量。用分光光度法和直接计数法分别测定M17培养基双麦麦精添加量为9.36g时,嗜热链球菌不同生长时间培养液的OD600值和菌体数量,绘制其生长曲线,并对2种测定方法所绘制的生长曲线进行比较,分析差异,见图5。
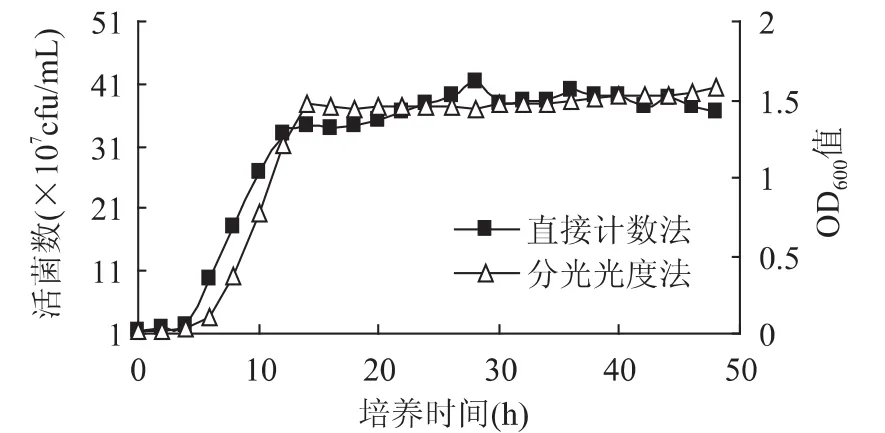
图5 嗜热链球菌生长曲线Fig.5 Growth curve of streptococcus thermophilus
两条曲线通过相关系数分析,相似度比较高,相似度系数为0.977。说明嗜热链球菌培养液OD600值的吸光度与直接计数细胞数量呈强相关性。分光光度法和直接计数法测定的生长曲线在对数期以前基本一致。在对数期,直接计数法所得生长曲线整体高于分光光度法所得生长曲线,可能由于菌体繁殖比较旺盛,使得菌体在培养基中成活率较高。稳定期分光光度法高于直接计数法,可能由于稳定期内菌体的自溶速度小于其缓慢的增长速度,同时随着代谢产物增多,使吸光度略微偏高。
2.3.2 嗜热链球菌在对数期的OD600值与直接计数法测得嗜热链球菌的关系 以OD600值为横坐标,直接计数法测定的细胞数量为纵坐标,绘制XY散点图,利用最小二乘法绘出两者的回归曲线,结果见图6。回归方程为y=-11.489x2+35.968x+6.1745(R2= 0.9998),回归方程具有极显著意义。
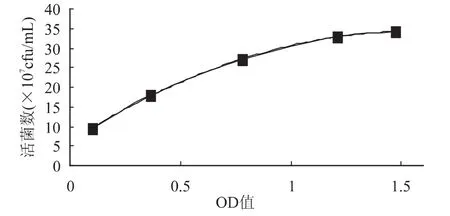
图6 对数生长期OD值与直接计数细胞数量的关系Fig.6 Relationship in logarithic growth phase between thr OD value and the number of cells calculated by direct counting method
3 结论与讨论
双麦麦精对嗜热链球菌的生长有明显的促进作用,可以为嗜热链球菌提供生长所需要的碳源和生长因子,但不能提供嗜热链球菌生长所需要的全部氮源,只能提供部分氮源。必须加入酵母膏、蛋白胨、牛肉膏等物质作为嗜热链球菌生长所需的氮源,这些物质中含有大量的氨基酸、肽类等可溶性含氮化合物、维生素和无机盐。这与万红兵等[18]的研究结果一致。嗜热链球菌可以利用葡萄糖、果糖、蔗糖、果糖作为碳源[19-20]。通过双麦麦精的添加,和乳糖同时存在,形成复合碳源,可以促进嗜热链球菌的生长,可以使嗜热链球菌能快速通过调整期和对数期,提早进入对数期,从而缩短整个培养时间。在有充足氮源的环境下,添加双麦麦精,可以明显的增加稳定期的最大菌数。
最终成分优化的结果为乳糖浓度5g/L,酵母膏浓度4g/L,蛋白胨浓度12g/L,牛肉膏浓度7g/L,双麦麦精浓度16g/L。为双麦麦精在M17培养基中的应用提供重要的依据。
用分光光度法和直接计数法分别绘制的嗜热链球菌生长曲线相似度较高。在严格控制实验条件的情况下,在对数期培养液的OD600值与实际菌体细胞数有强相关性,在对数生长期可通过测定培养液的OD600值计算实际嗜热链球菌活菌数,也可以根据培养液的OD600值判断菌体生长是否进入对数期。
[1]王建芳,陈芳,靳亚平.嗜热链球菌适宜培养条件研究[J].西北农业学报,2008,17(2):56-58.
[2]Iyer R,Tomar S K,Maheswari T U,et al.Streptococcus Thermophilus Strains:Multifunctional Lactic Acid Bacteria[J].International Dairy Journal,2010,20(3):133-141.
[3]黄小丹,柳建良,杜林.酸乳中乳酸菌计数培养基的优选与改良[J].仲恺农业技术学院学报,2006,19(1):21-24.
[4]辛若竹,丁梅,孟宪志,等.活性酸乳中每种益生菌总数的同时计数法[J].食品科学,2007,28(1):244-248.
[5]陈芳,靳亚平.嗜热链球菌部分生物学特性研究[J].中国奶牛,2007(8):42-44.
[6]Wouters J T M,Ayad E H E,Hugenholtz J,et al.Microbes from Raw Milk for Fermented Dairy Products[J].International Dairy Journal,2002,12(2-3):91-109.
[7]李莹,周剑忠,黄开红.嗜热链球菌蔬菜汁培养基及培养条件的优化[J].食品工业科技,2007,28(10):148-150.
[8]Valduga E,Tatsch P O,Tiggemann L,et al.Evaluation of the Conditions of Carotenoids Production in a Synthetic Medium by Sporidiobolus Salmonicolor(Cbs 2636)in a Bioreactor[J].International Journal of Food Science and Technology,2009,44 (12):2445-2451.
[9]Valduga E,Valerio A,Treichel H,et al.Optimization of the Production of Total Carotenoids by Sporidiobolus Salmonicolor (Cbs 2636)Using Response Surface Technique[J].Food and Bioprocess Technology,2009,2(4):415-421.
[10]陈如琪,吕宝龙,卢艳花,等.双麦麦精对酸奶益生菌及其感官性质的影响[J].中国乳品工业,2012,40(3):39-42.
[11]Aly M E.Malt extract for enhancing flavour development of ultrafiltered domiati cheese[J].Food Chemistry,1997,59(1): 127-133.
[12]Liu Q,Qiu Y,Beta T.Comparison of antioxidant activities of different colored wheat grains and analysis of phenolic compounds[J].Journal of Agricultural and Food Chemistry,2010,58(16): 9235-9241.
[13]陈龄,丁文平,曹银.液体大麦精在面包烘焙中的应用[J].中国粮油学报,2009,24(7):18-22.
[14]古元懿,欧军,梁金钟.保加利亚乳杆菌和嗜热链球菌促生长因子的研究[J].乳业科学与技术,2008,31(3):117-120.
[15]古元懿,欧军,梁金钟.保加利亚乳杆菌和嗜热链球菌促生长剂的研究[J].中国奶牛,2008(7):44-47.
[16]Patel H M,Pandiella S S,Wang R H,et al.Influence of malt, wheat,and barley extracts on the bile tolerance of selected strains of lactobacilli[J].Food Microbiology,2004,21(1):83-89.
[17]Pyar H,Peh K K.Effect of cryoprotective agents on survival and stability of lactobacillus acidophilus cultured in food-grade medium[J].International Journal of Dairy Technology,2011,64 (4):578-584.
[18]万红兵,田洪涛,山丽杰,等.嗜热链球菌与保加利亚乳杆菌麦芽复合汁增菌培养基的优化筛选[J].食品与发酵工业,2006,32(6):51-55.
[19]刘丽波,李春,孟祥晨.1株高产eps嗜热链球菌的筛选及培养条件优化[J].中国食品学报,2007,7(3):64-68.
[20]Van Den Bogaard P T C,Hols P,Kuipers O P,et al.Sugar Utilisation and Conservation of the Gal-Lac Gene Cluster in Streptococcus Thermophilus[J].Systematic and Applied Microbiology,2004,27(1):10-17.
Effect of ABF Bio+malt extract on the growth ofStreptococcus thermophilus
ZHAO Shuang1,LIU Shao-wei1,*,CHEN Ru-qi1,LV Bao-long1,LU Yan-hua1,FEI Nan2
(1.East China University of Science and Technology,Shanghai 200237,China; 2.Shanghai AB Food&Beverages Ltd.,Shanghai 200245,China)
The effects of ABF Bio+malt extract on the main nutrition constituent of theStreptococcus thermophilesin its growth progress by adding ABF Bio+malt in the incomplete nutrient medium were studied.The number of viable cells inStreptococcus thermophilescounted according to its Bacilli absorbance was used to analyse the effect onStreptococcus thermophilesbrought by ABF Bio+malt extract.The optimal proportion of ABF Bio+malt extract and other nutrients was determined by orthogonal experiment with direct counting method.The results showed that ABF Bio+malt extract played an important role of the augmentor the growth ofstreptococcus thermophilusas well as a provider of sufficient carbon source and growth factors for it.The ABF Bio+malt extract produced the best result when its additive amount reached 16g/L.After analysis and comparison of the growth curves which were measured by absorb value method and the direct counting method respectively,the curves were proved to be of good similarity,especially in the logarithmic phase.
Streptococcus thermophilus;growth curve;ABF Bio+malt extract;default medium;colony counts
TS201.3
A
1002-0306(2012)21-0062-05
2012-04-10 *通讯联系人
赵爽(1990-),男,本科,研究方向:食品科学。
中央高校基本科研业务费专项资金;上海科委资助课题(10dz1200904)。
