英夫利昔、沙利度胺对大鼠实验性结肠炎的治疗作用及其机制研究*
2012-10-22盛露露杨晓娣朱水津马天乐江石湖
盛露露 杨晓娣 朱水津 马天乐 江石湖
上海交通大学医学院附属瑞金医院消化内科(200025)
炎症性肠病(IBD)包括溃疡性结肠炎(UC)和克罗恩病(CD),欧美国家比较常见,近年发展中国家的发病率呈上升趋势[1]。CD是一种慢性非特异性、复发与缓解交替的肉芽肿性炎症,目前病因尚未明确,缺乏根治药物,需长期维持用药,传统疗法存在疗效不佳和安全性问题。随着对IBD发病机制的深入研究,其治疗已进入免疫调节分子靶向治疗的时代。生物制剂英夫利昔是针对肿瘤坏死因子-α(TNF-α)的单克隆抗体,临床效果明显且见效快,但其具有选择性,且需序列重复用药,价格昂贵,长期输注的安全性亦有待证实[2,3]。近年研究表明沙利度胺对TNF-α mRNA具有降解作用,对IBD的疗效基本得到肯定。且相对英夫利昔,沙利度胺口服方便、价格低,但应注意其可能的毒副作用[4]。本实验拟通过制备三硝基苯磺酸(TNBS)大鼠结肠炎模型,并比较英夫利昔、沙利度胺的治疗效果,旨在从免疫调节、抑制病理性血管生成、抑制凋亡三方面探讨两者治疗IBD的作用机制。
材料与方法
一、材料
1.实验动物:健康成年雄性Sprague-Dawley(SD)大鼠 46 只,体质量(200±10) g,均购自中国科学院上海生命科学研究院。饲养条件均为SPF级,昼夜节律12 h/12 h,无特殊实验要求时正常供应饲料和饮水。购入后适应性饲养1周。
2.主要试剂和仪器:50 g/L TNBS溶液(Sigma-Aldrich Co.);英夫利昔(RemicadeTM,美国 Centocor公司惠赠);沙利度胺(商品名:反应停,常州制药厂有限公司);TRIZOL抽提试剂(Invitrogen by Life Technologies);普通PCR试剂盒、逆转录试剂盒、Real-time PCR 试剂盒(TaKaRa Bio Inc.);质粒抽提试剂盒[生工生物工程(上海)有限公司];兔抗大鼠 TNF-α、血管内皮生长因子(VEGF)、caspase-3 多克隆抗体(Abcam plc.);GAPDH 多克隆抗体、HRP标记的山羊抗兔二抗(上海康成生物工程有限公司);TUNEL细胞凋亡检测试剂盒(南京凯基生物科技发展有限公司);免疫组化试剂盒(福州迈新生物技术开发有限公司)。实时定量PCR仪(Stratagene Mx3000P QPCR System,Agilent Technologies,Inc.);普通 PCR 仪(GeneAmp,PCR system 9700);凝胶成像分析系统(美国Bio-Rad);紫外分光光度计(Pharmacia Biotech,Ultrospec 3000)。
二、实验方法
1.结肠炎模型的建立和药物干预:大鼠随机分为正常对照组(n=10)、结肠炎组(n=12)、英夫利昔组(n=12)和沙利度胺组(n=12)。造模前大鼠禁食不禁水36 h,经乙醚麻醉后,将直径2 mm的硅胶管从肛门插入肠道深约8 cm处,注入含TNBS 20 mg的50%(v/v)乙醇溶液约 0.8 mL(TNBS 按照 100 mg/kg给药加上等体积的无水乙醇)[5];正常组给予等量0.9%NaCl溶液灌肠,倒悬位持续5 min。造模后第1 d,观察各组大鼠一般情况,将制模成功的大鼠纳入后续实验,每天同一时间段给予药物干预,英夫利昔组:英夫利昔(10 mg/mL)腹腔注射 5 mg/kg[6];沙利度胺组:沙利度胺药片碾磨成粉状后溶于橄榄油(50 mg/mL)经管喂,200 mg/kg[7];正常对照组/结肠炎组:每组各一半大鼠分别给予等量橄榄油管喂或0.9%NaCl溶液腹腔注射,连续治疗7 d。
2.观察指标:①大鼠的一般情况:每天固定时间段内观察大鼠精神状态、毛发光泽、活动、饮食情况等并记录体质量变化、大便性状和便血情况,对后三项行疾病活动指数(DAI)评分[8]。②大体形态损伤指数(CMDI)评分:连续给药7 d后,处死各组大鼠,取远端结肠约12 cm,冲洗干净后行局部结肠CMDI评分[9]。 ③组织损伤指数(TDI)评分:取部分病变肠段组织立即放于40 g/L多聚甲醛液中固定,石蜡包埋后切片行HE染色和TDI评分[10]。
3.Real-time PCR检测大鼠结肠组织TNF-α、VEGF、caspase-3 mRNA表达:取适量结肠组织匀浆后,按TRIZOL说明书抽提总RNA,逆转录成cDNA,行PCR扩增(PCR特异性引物序列见表1)。PCR反应产物电泳鉴定后割胶回收纯化。PMD-18T载体连接PCR纯化产物,连接产物转化感受态细菌,继而行细菌扩增和培养。挑选克隆行PCR鉴定,选取条带与目的条带相符的细菌行基因测序。测序结果与Genbank相应序列匹配一致后,抽提菌液质粒,计算基因浓度。将已知基因浓度的质粒逐步10倍稀释后,行Real-time PCR反应。绘制各个目的基因和管家基因的标准曲线。将各组结肠样本cDNA行Real-time PCR反应,对照标准曲线,计算各标本中不同基因的拷贝数。
4.蛋白质印迹法检测 TNF-α、VEGF、caspase-3蛋白表达:取适量结肠组织加入1 mL RIPA匀浆,冰浴30 min后12000 r/min 4℃离心30 min。取上清水煮变性后行SDS-PAGE电泳,然后转移至硝酸纤维素膜,电压100 V,时间1 h 30 min,5%脱脂奶粉 37℃封闭 1 h;分别加入兔抗大鼠 TNF-α、VEGF、caspase-3 多克隆抗体(1∶1000 稀释),4 ℃孵育过夜;加入二抗(1∶3000 稀释),室温孵育 1 h,ECL显影;Image-Pro Plus 6.0软件行灰度扫描分析蛋白表达的相对值。

表1 PCR引物序列
5.TUNEL法检测结肠上皮细胞凋亡:常规脱蜡水化,20 g/L蛋白酶K 37℃消化30 min,PBS漂洗后浸入封闭液(3%H2O2溶于甲醇),室温封闭10 min,加 rTdT 酶反应液反应 60 min,以 1∶100 稀释的 Streptavidin-HRP 37℃反应 30 min,DAB显色,苏木精复染,封片镜检。细胞呈棕褐色颗粒者,并结合细胞形态学改变,判断为凋亡细胞。
6.免疫组化法检测 TNF-α、VEGF、caspase-3 表达:常规脱蜡至水;3%H2O2室温孵育10 min,灭活内源性过氧化物酶活性;正常山羊血清室温封闭 20 min;滴加兔抗大鼠 TNF-α(1∶2000)、VEGF(1∶200)、caspase-3(1∶100)一抗,4 ℃过夜;加入山羊抗兔二抗室温静置或37℃ 1 h;DAB显色5~10 min,苏木精复染、脱水、透明、封片、镜检。
三、统计学分析
结 果
一、结肠炎模型
1.一般情况:造模后24 h,正常对照组大鼠一般情况基本正常,体质量基本无变化。其余三组大鼠均出现稀便、血便或隐血试验阳性,毛发竖起,精神不佳,饮食、活动减少,体质量下降。结肠炎组造模后第2~4 d平均DAI评分达到高峰,在造模后7 d内无明显下降趋势;而英夫利昔组、沙利度胺组给药后第2 d开始部分大鼠一般情况稍有改善,稀便、便血减少,活动、饮食增加,同时体质量开始缓慢升高。结肠炎组平均DAI评分明显高于正常对照组(P<0.01),英夫利昔组、沙利度胺组明显低于结肠炎组(P<0.01),英夫利昔组平均DAI评分明显低于沙利度胺组(P<0.01)(见表 2)。
2.大体形态学改变:正常对照组大鼠肠壁薄而光滑,周围结构清楚,无黏连。结肠炎组大鼠病变部位肠黏膜充血水肿,严重节段性糜烂、溃疡、坏死,肠壁与周围组织黏连,肠腔狭窄,部分大鼠结肠近端呈巨结肠样改变,肠壁僵硬易断;英夫利昔组、沙利度胺组仅表现为局部黏膜充血水肿,糜烂溃疡基本愈合,肠壁增厚或瘢痕形成,黏连无或减轻,少部分存在轻度糜烂和浅溃疡。结肠炎组CMDI评分明显高于正常对照组(P<0.05),英夫利昔组、沙利度胺组明显低于结肠炎组(P<0.05),英夫利昔组与沙利度胺组相比无明显差异(P>0.05)(见表2)。
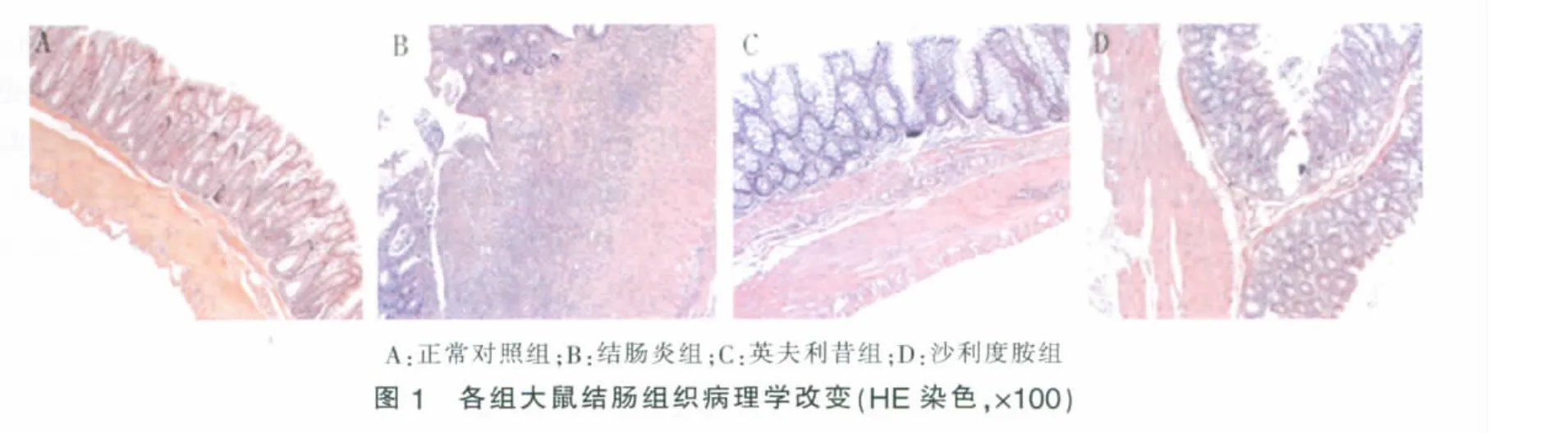
3.组织病理学改变:正常对照组结肠上皮完整,细胞形态正常,固有层腺体排列规则,各层结构清晰;结肠炎组黏膜上皮坏死脱落,溃疡糜烂形成,全层透壁性炎症,大量中性粒细胞、淋巴细胞以及少量嗜酸性粒细胞浸润,可见血管增生和肉芽组织,全层结构消失;英夫利昔组、沙利度胺组溃疡基本愈合,炎性细胞浸润减少,肠壁结构基本恢复(见图1)。结肠炎组平均TDI评分明显高于正常对照组(P<0.05),英夫利昔组、沙利度胺组明显低于结肠炎组(P<0.05),两治疗组无明显差异(P>0.05)(见表 2)。
表2 各组大鼠平均 DAI、CMDI、TDI评分比较()

表2 各组大鼠平均 DAI、CMDI、TDI评分比较()
与正常对照组比较,*P<0.05,**P<0.01;与结肠炎组比较,▲P<0.05,▲▲P<0.01;#与沙利度胺组比较,P<0.01
二、Real-time PCR结果
结肠炎组结肠组织 TNF-α、VEGF、caspase-3 mRNA表达均明显高于正常对照组(P<0.05),英夫利昔组、沙利度胺组明显低于结肠炎组(P<0.05),而两治疗组间均无明显差异(P>0.05)(见表3)。
表3 各组大鼠结肠组织TNF-α、VEGF、caspase-3 mRNA表达比较()

表3 各组大鼠结肠组织TNF-α、VEGF、caspase-3 mRNA表达比较()
*与正常对照组比较,P<0.05;#与结肠炎组比较,P<0.05
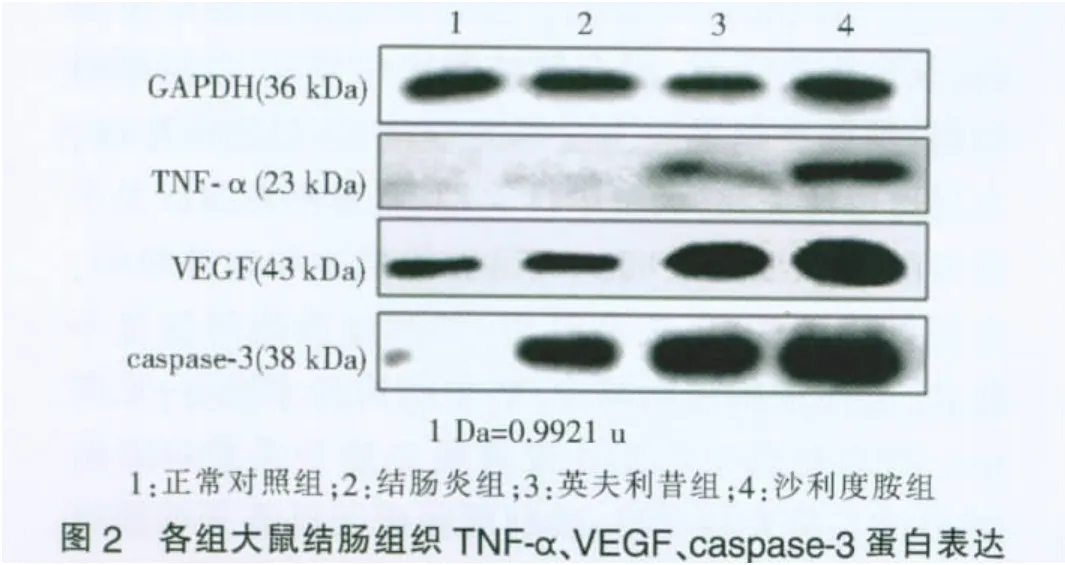
三、蛋白质印迹结果
与正常对照组相比,结肠炎组结肠组织TNF-α、VEGF、caspase-3 蛋白表达均明显升高(P<0.05),英夫利昔组、沙利度胺组蛋白表达较结肠炎组均明显下降(P<0.05),两治疗组间TNF-α蛋白表达无明显差异,而英夫利昔组VEGF、caspase-3蛋白表达均明显低于沙利度胺组(P<0.05)(见图2)。
四、结肠上皮细胞凋亡
正常对照组大鼠结肠上皮仅少量凋亡细胞,结肠炎组凋亡现象普遍,英夫利昔组、沙利度胺组大鼠结肠上皮细胞凋亡明显改善(见图 3)。
五、免疫组化
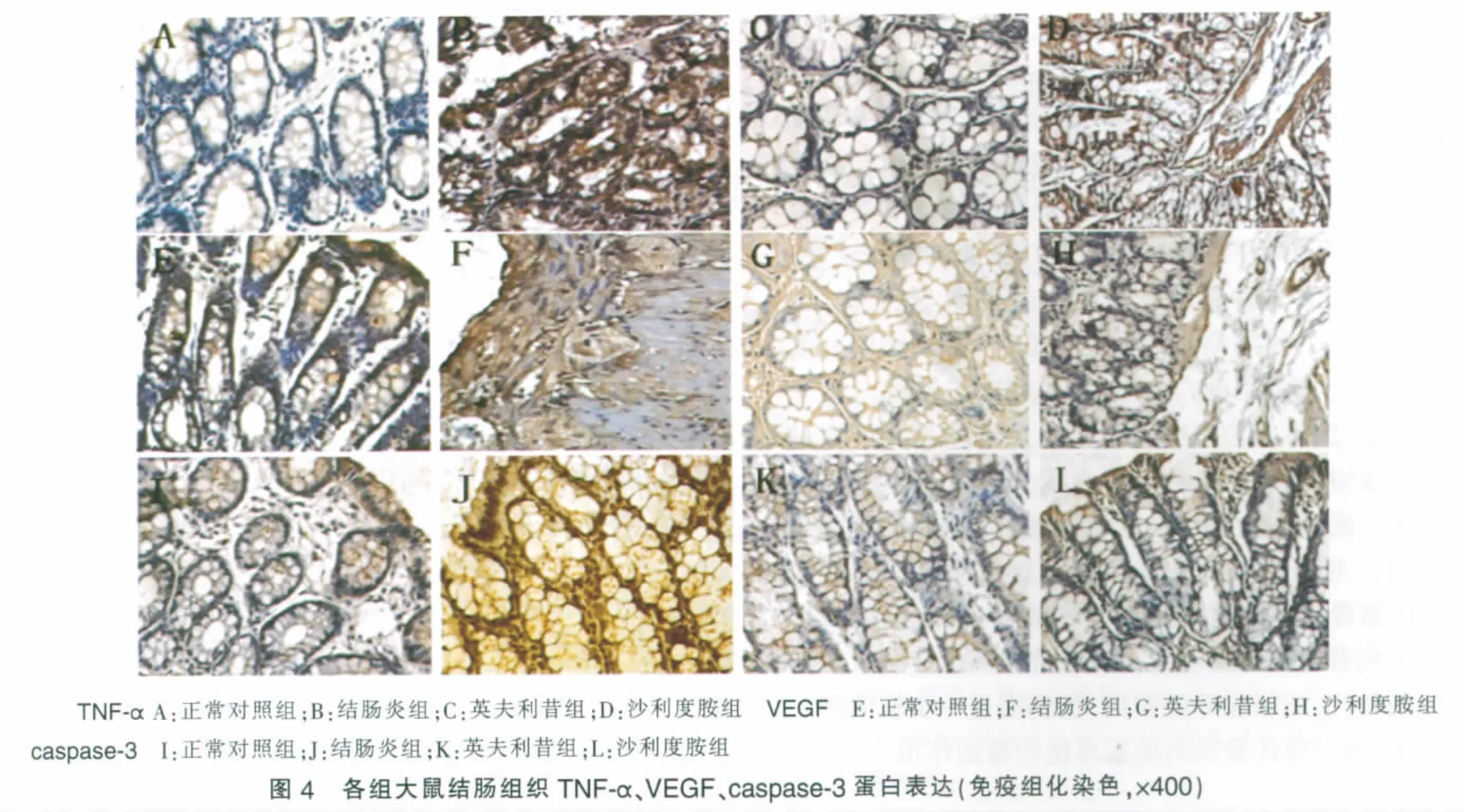
TNF-α在正常对照组大鼠肠组织中基本无表达,结肠炎组肠上皮细胞质中可见明显棕色强表达,而英夫利昔组、沙利度胺组TNF-α表达明显减弱,呈散在表达;VEGF在正常对照组表达较弱,结肠炎组肠上皮细胞、血管内皮细胞及其间质炎细胞、成纤维细胞中均可见VEGF强表达,英夫利昔组、沙利度胺组表达均明显减弱;正常对照组caspase-3表达较弱,结肠炎组表达显著增强,阳性细胞呈棕黄色,主要位于肠上皮细胞质和部分胞核,英夫利昔组、沙利度胺组表达明显减弱(见图4)。
讨 论
目前IBD的发病机制尚不清楚,建立适当的肠道炎症动物模型对研究其发病机制和药物疗效具有积极意义。在多种构建肠道炎症动物模型的方法中,TNBS/乙醇法目前的应用最为广泛,该方法诱发的体内免疫反应以Th1型反应为主,类似于人类CD。本实验采用TNBS/乙醇混合液灌肠制备大鼠结肠炎模型,结肠炎组大鼠平均DAI、CMDI、TDI评分与正常对照组相比均显著升高,提示造模成功。
肠道黏膜免疫异常在IBD的发病中处于中心地位,反复抗原刺激使肠黏膜免疫细胞聚积、激活,并刺激黏膜固有层细胞因子激活和分泌增加,如白细胞介素(IL)-6、IL-12、TNF-α、干扰素(IFN)-γ 等,促进Th1型炎症的发生。TNF-α作为一种促炎症因子和免疫调节因子,在CD免疫反应和病理过程中起关键作用。研究表明活动性IBD患者血液、肠上皮组织以及粪便中TNF-α水平均明显升高[11]。

IBD患者肠黏膜组织反复形成溃疡和上皮再生,血管生成是IBD中的必然过程。正常情况下,促血管形成因子与抑制因子处于平衡状态,一旦打破会激活血管系统,使血管过度生成或抑制。促血管形成因子-抑制因子的平衡状态、微环境以及IBD患者异常表达的细胞因子,可调节生理性血管生成向病理性血管生成的演变,且这种病理性血管生成可能对IBD慢性炎症的发生和维持起一定作用。近年有学者提出病理性血管生成可作为IBD的发病因素之一[12,13]。另一方面,炎症和免疫反应对病理性血管生成亦起调节作用。有研究[14]发现,在IL-10基因敲除和DSS诱导的小鼠慢性和急性肠道炎症模型中,仅在活动性炎症浸润的区域可见明显血管生成。VEGF是最直接的血管内皮细胞促分裂素,与受体结合后发挥一系列促进血管生成的生物学功能。研究[15]表明血清VEGF水平与IBD的疾病活动度密切相关,VEGF是监测血管生成的重要指标之一。
目前,越来越多的研究证实IBD炎症肠黏膜组织内存在细胞凋亡紊乱。肠上皮细胞是肠黏膜屏障的重要机械屏障,完整的肠上皮细胞不仅能阻止细菌和毒素等大分子渗透至肠黏膜固有层,还能激活固有层免疫细胞以维护肠黏膜屏障功能的稳定,避免黏膜异常免疫反应的发生[16]。研究表明IBD存在大量结肠上皮细胞凋亡,导致上皮损伤,破坏肠黏膜屏障,这也是造成IBD肠道炎症发生和持续的重要原因[17]。caspase是一类与凋亡密切相关的蛋白水解酶家族,caspase-3在凋亡的执行阶段对全部或部分关键性蛋白进行酶切,从而引起凋亡的激活或灭活,活化的caspase-3可作为细胞凋亡的标记。
慢性复发顽固性IBD患者的治疗仍是临床面临的挑战,高达1/3的IBD患者对传统治疗无效或耐受。对于这部分患者,英夫利昔单抗的疗效较好。英夫利昔是针对TNF-α的人鼠嵌合性单克隆抗体,与TNF-α结合后可激活补体和抗体介导的活动性CD4+T细胞和巨噬细胞的细胞毒反应。目前英夫利昔已用于对常规保守治疗无效以及活动性瘘管形成的中重度CD患者。虽然生物治疗的疗效肯定,但部分患者的身体状况以及经济情况限制了其使用,且随着抗抗体产生而降低疗效,生物制剂长期使用的安全性问题(如感染、恶性肿瘤等)亦存在诸多争议[18]。沙利度胺具有抑制TNF-α的作用,近年发现其具有抗炎、免疫调节、抑制血管生成等活性。1997年Wettstein等[19]首先采用沙利度胺治疗1例对皮质类固醇治疗无效的CD患者,取得良好效果。动物模型研究[20]发现沙利度胺通过下调促炎因子基因的表达而损伤内皮细胞-粒细胞的相互作用,对TNBS诱导的CD样结肠炎起治疗作用;Bariol等[21]发现沙利度胺对IBD患者症状的短期疗效较好;罗政仁等[22]的随机对照研究认为沙利度胺对轻中度CD患者有效,且其疗效优于传统药物柳氮磺吡啶。由此可见,沙利度胺可望成为治疗IBD的一种新型药物。但沙利度胺治疗CD的作用机制较复杂,尚未有统一的定论。有学者[23]指出对英夫利昔治疗有反应的患者,沙利度胺可作为其维持缓解的桥接治疗。
本实验中,结肠炎组 DAI、CMDI、TDI评分明显升高,TNF-α、VEGF、caspase-3 mRNA 和蛋白表达亦显著升高,肠上皮细胞凋亡显著增加;英夫利昔组、沙利度胺组上述各项指标均较结肠炎组明显改善,且英夫利昔组VEGF蛋白表达明显低于沙利度胺组,提示两种治疗药物对IBD炎症的疗效较好;能有效抑制IBD病理性血管生成,且英夫利昔抑制血管生成的作用略优于沙利度胺;两种药物能有效抑制肠上皮细胞凋亡,保护肠黏膜上皮屏障。
总之,本实验表明英夫利昔、沙利度胺均能通过抑制TNF-α、VEGF、caspase-3表达, 对 IBD 的免疫失衡、病理性血管生成以及肠上皮细胞凋亡起调节作用。对于传统治疗无效且无法使用英夫利昔的IBD患者,沙利度胺可能是治疗的一种新选择;对于英夫利昔治疗有反应者,其维持缓解治疗可以沙利度胺替代,能明显降低费用。但仍需进一步临床研究验证,同时应注意沙利度胺可能的毒副作用。
1 Bernstein CN,Shanahan F.Disorders of a modern lifestyle:reconciling the epidemiology of inflammatory bowel diseases[J].Gut,2008,57(9):1185-1191.
2 Caspersen S,Elkjaer M,Riis L,et al.Infliximab for inflammatory bowel disease in Denmark 1999-2005:clinical outcome and follow-up evaluation of malignancy and mortality[J].Clin Gastroenterol Hepatol,2008,6(11):1212-1217.
3 Stallmach A,Hagel S,Bruns T.Adverse effects of biologics used for treating IBD[J].Best Pract Res Clin Gastroenterol,2010,24(2):167-182.
4 Ng SC,Chan FK,Sung JJ.Review article:the role of nonbiological drugs in refractory inflammatory bowel disease[J].Aliment Pharmacol Ther,2011,33(4):417-427.
5 Tirosh O,Levy E,Reifen R.High selenium diet protects against TNBS-induced acute inflammation,mitochondrial dysfunction,and secondary necrosis in rat colon[J].Nutrition,2007,23(11-12):878-886.
6 Fries W,Muja C,Crisafulli C,et al.Infliximab and etanercept are equally effective in reducing enterocyte APOPTOSIS in experimental colitis[J].Int J Med Sci,2008,5(4):169-180.
7 Carvalho AT,Souza H,Carneiro AJ,et al.Therapeutic and prophylactic thalidomide in TNBS-induced colitis:synergisticeffectson TNF-alpha,IL-12 and VEGF production[J].World J Gastroenterol,2007,13(15):2166-2173.
8 Murano M,Maemura K,Hirata I,et al.Therapeutic effect of intracolonically administered nuclear factor kappa B(p65)antisense oligonucleotide on mouse dextran sulphate sodium(DSS)-induced colitis[J].Clin Exp Immunol,2000,120(1):51-58.
9 Butzner JD,Parmar R,Bell CJ,et al.Butyrate enema therapy stimulates mucosal repair in experimental colitis in the rat[J].Gut,1996,38(4):568-573.
10 王皓,欧阳钦,胡仁伟.三硝基苯磺酸结肠炎动物模型的建立[J].胃肠病学,2001,6(1):7-10.
11 张子其,刘婧.肿瘤坏死因子与炎症性肠病[J].中国实用内科杂志,2007,27(18):1439-1442.
12 Danese S,Sans M,de la Motte C,et al.Angiogenesis as a novel component of inflammatory bowel disease pathogenesis[J].Gastroenterology,2006,130(7):2060-2073.
13 Pousa ID,Gisbert JP,Maté J.Vascular development in inflammatory bowel disease (Article in Spanish)[J].Gastroenterol Hepatol,2006,29(7):414-421.
14 Danese S,Sans M,Spencer DM,et al.Angiogenesis blockade as a new therapeutic approach to experimental colitis[J].Gut,2007,56(6):855-862.
15 Kapsoritakis A,Sfiridaki A,Maltezos E,et al.Vascular endothelial growth factor in inflammatory bowel disease[J].Int J Colorectal Dis,2003,18(5):418-422.
16 毛靖伟,王英德.肠黏膜屏障在炎症性肠病中作用机制的研究进展[J].世界华人消杂志,2010,18(7):695-698.
17 Salim SY,Söderholm JD.Importance ofdisrupted intestinal barrier in inflammatory bowel diseases[J].Inflamm Bowel Dis,2011,17(1):362-381.
18 顾于蓓,袁耀宗.生物制剂治疗在炎症性肠病中的长期疗效和安全性问题[J].中华消化杂志,2010,30(1):63-66.
19 Wettstein AR,Meagher AP.Thalidomide in Crohn’s disease[J].Lancet,1997,350(9089):1445-1446.
20 Lienenlüke B,Stojanovic T,Fiebig T,et al.Thalidomide impairment of trinitrobenzene sulphonic acid-induced colitis in the rat-role of endothelial cell-leukocyte interaction[J].Br J Pharmacol,2001,133(8):1414-1423.
21 Bariol C,Meagher AP,Vickers CR,et al.Early studies on the safety and efficacy of thalidomide for symptomatic inflammatory bowel disease[J].J Gastroenterol Hepatol,2002,17(2):135-139.
22 罗政仁,姒建敏.沙利度胺治疗克罗恩病疗效观察[J].现代消化及介入诊疗,2008,13(2):82-83.
23 Sabate JM,Villarejo J,Lemann M,et al.An open-label study of thalidomide for maintenance therapy in responders to infliximab in chronically active and fistulizing refractory Crohn’s disease[J].Aliment Pharmacol Ther,2002,16(6):1117-1124.
