水稻新资源温敏核不育系长S育性基因的等位性测验
2012-10-21张蓓玲王安萍段世华许东风郑自伟
郑 卓,张蓓玲,王安萍 ,吴 杨 ,贺 俐,段世华,许东风 ,郑自伟
水稻新资源温敏核不育系长S育性基因的等位性测验
*郑 卓1,张蓓玲2,王安萍1,吴 杨1,贺 俐1,段世华1,许东风1,郑自伟1
(1.井冈山大学生命科学学院,江西,吉安 343009;2.吉安市农业局,江西,吉安 343000)
长S来自普通野生稻与籼稻珍珠矮杂交后代。在自然条件下,长S表现为长日高温可育、低温不育,且育性转换明显。经等位性测验,长S与C815S、广占63S、株1S、湘陵628S及HD9802S的育性基因等位,而与HN5S、培矮64S的育性基因不等位。
水稻;温敏核不育;长S;等位性测验
自1973年石明松[1]在晚粳品种农垦58群体中发现光敏感核不育株后,我国开始了两系法杂交水稻的研究。经过近四十年的研究,我国在两系法杂交水稻研究上取得了成功并且在生产上得到了大面积的利用。截止2005年,通过省级鉴定(审定)的光温敏核不育系106个;1994~2009年通过省级和国家审定的两系杂交水稻组合427个;2009年我国两系杂交水稻播种面积已达到杂交水稻总播种面积的20.9%[2]。但目前这些两系杂交水稻的亲本主要集中在广占63S及其衍生系、安农S-1衍生系以及株1S、陆18S和HD9802S上[3]。不育系来源单一,严重制约了我国两系杂交水稻的进一步发展。因此,积极发掘光温敏核不育新资源,丰富两系不育系的来源,扩大遗传基础,这对保证我国两系杂交水稻的可持续发展具有十分重要的意义。
长S系笔者在广西普通野生稻()与珍珠矮(Kato)杂交后代中发现的新资源温敏核不育材料,具有穗大粒多等特点[4]。长S的获得丰富了我国两系不育系的来源,扩大了遗传基础,但长S的不育基因与现有温敏核不育系的不育基因是否等位尚不清楚。本文对这一问题进行了研究,旨在为长S的转育及利用提供参考。
1 材料与方法
1.1 试验材料
被测系长S由井冈山大学水稻繁育研究中心提供,籼型光温敏核不育系HN5S由长江大学农学院戴绍钧教授提供,培矮64S、广占63S由国家杂交水稻工程技术研究中心何强提供,C815S由湖南农业大学水稻研究所陈立云研究员提供,籼型温敏核不育系株1S、湘陵628S由湖南亚华种业杨远柱研究员提供,HD9802S由湖北省农科院水稻研究所武晓智提供。各不育系的系谱来源、不育基因对数及性质[4-17]如表1。
1.2 试验方法
试验于2010~2011年在井冈山大学生命科学学院试验基地(江西·吉安,N 27°07′)进行。
1.2.1 自然条件下长S育性变化观察
2011年在井冈山大学试验田对长S进行分期播种,第1期播种时间为4月24日,以后每7d播种1期,共播种10期。每期长S的栽插规格为3行×10列,栽插密度为20.0 cm × 20.0 cm,栽培管理同大田。育性检查从第1期播种材料7月5日见穗开始至9月下旬结束,每3 d镜检1次。每次取3株,每株取1穗镜检花粉育性,每穗上、中、下部各取成熟颖花1~2朵,剥取花药,用镊子将花药在载玻片上捣碎,用1%I-IK溶液染色,置于40×显微镜下观察,每片观察3个视野,要求花粉较分散易计数,且每个视野花粉数在150粒以上,计数可染花粉的百分率。花粉区分为败育(包括典败和圆败)和可育(染色)两类。计算可染花粉率。以花粉可染率作为育性表现的直观分析。光长参照参考文献[18]的旬平均日长,温度资料来自吉安市气象局2011年气象资料。可染花粉率的计算公式如下:
可染花粉率=可染花粉数/总花粉数×100%

表1 8个光(温)敏核不育系的系谱来源、不育基因对数及性质
*广占63S虽衍生自农垦58S,但其不育基因的性质目前还没明确。
1.2.2 不育基因等位性测验
2011年2月下旬在海南三亚低温条件下, HN5S、C815S、广占63S、湘陵628S和HD9802S可育期内,以长S为母本,分别与这7个光(温)敏核不育系杂交,获得杂种F1种子。2011年将这7个杂种F1和8个光温敏核不育系作中稻正季种植,以保证其小孢子分化均处于高温和长日照条件下。5月1日播种,小区5行×10列,栽插密度20.0 cm × 20.0 cm,栽培管理同大田。杂种F1及各不育系在始穗期随机选取3个单株,每个单株选取3个单穗套袋自交,并镜检单株花粉育性,方法同上。成熟后考查套袋自交结实率,计算公式如下:
套袋结实率% =(小穗实粒数/小穗总粒数)×100%
2 结果与分析
2.1 长S在不同自然光长、温度条件下的育性表现
2011年对长S在不同自然光长、温度条件下的育性表现进行了观察,结果如表2。在吉安自然条件下,长S自7月5日见穗至9月6日均处于完全不育阶段,9月6日后发生了育性转换,这一结果说明长S的育性受自然光长、温度条件的控制,主要表现为长日高温不育,低温可育。8月1~8日连续8 d吉安出现了低温天气,其中8月1日、3日、8日最低气温为24 ℃,8月2日、4日、5日为23 ℃, 8月6日为20 ℃,8月7日为22 ℃,长S遭遇这一低温后仍保持高度不育。这一结果说明在长日条件下长S的不育起点温度可能较低,上述低温没能达到长S的不育起点温度,或者是虽低温达到了长S的不育起点温度,但低温持续的时间较短,没能达到一定的低温积累量,因而长S的育性没有发生波动,仍表现为高度不育。8月20日后吉安出现了日最低气温达24℃以下的持续低温天气,导致了9月中下旬抽穗的长S育性发生了明显转换。这一结果说明在短日条件下长S的不育起点温度可能升高,上述低温达到了长S的不育起点温度,而且低温持续的时间较长,达到了一定的低温积累量,因而长S的育性发生了转换,表现为可育。综合上述结果,在一定的日照长度条件下,要使温敏核不育系长S的育性发生变化,必须达到一定的低温程度和低温持续时间。

表2 长S分期播种育性表现(江西·吉安)
*1)表示长S已进入幼穗分化;
2.2 光(温)敏核不育系不育期育性表现
长S等8个不育系均于2011年5月1日播种,始穗期在7月10至8月1日,始穗后调查这8个不育系的育性表现,结果如表3。由表中结果可知,长S与其它7个不育系在长日高温自然条件下花粉可染率和套袋结实率均为0,不育系的不育性表现正常。长S的生育期与早籼型温敏核不育系HD9802S一致,属于早籼类型的两用核不育系。
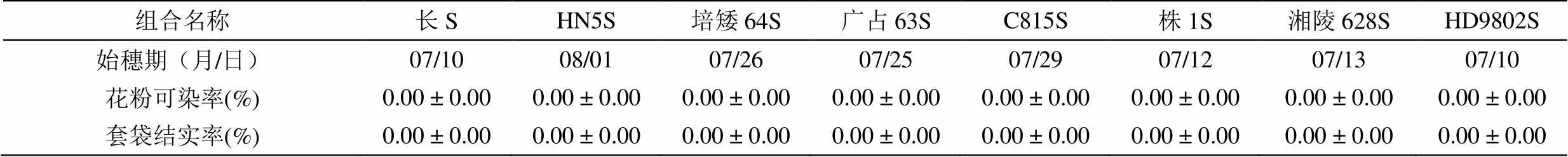
表3 长S与其它光(温)敏核不育系不育敏感期育性表现
2.3 各不育系与长S杂交F1代育性表现
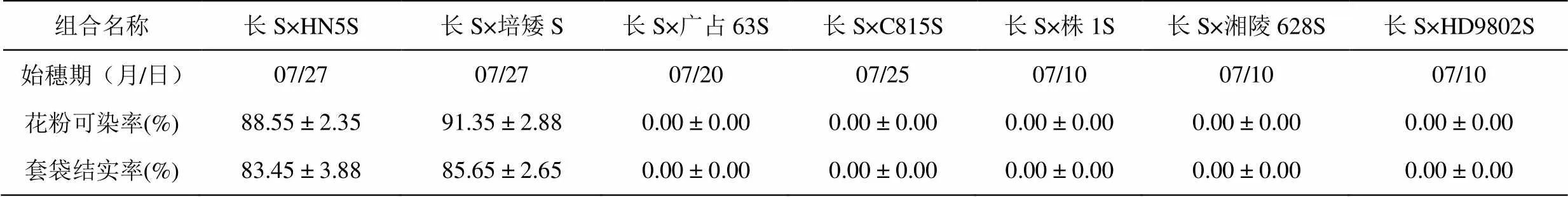
表4 长S与7个光(温)敏核不育系杂交F1的育性表现(江西·吉安)
长S与HN5S、广占63S等杂种F1均在2011年7月10日~27日始穗,始穗期均在不育系稳定不育期内,考查杂种F1的育性表现,结果列于表4。由表中结果可知,长S与HN5S、培矮64S杂种F1的花粉育性和套袋结实率均表现为正常可育,表明长S与HN5S、培矮64S的不育基因是非等位的;而长S与广占63S、C815S、株1S、湘陵628S、HD9802S杂种F1的花粉育性和套袋结实率均为0,表现为不育,表明长S与广占63S、C815S、株1S、湘陵628S、HD9802S的不育基因是等位的。周勇等[17]研究表明HD9802S的育性受一对隐性核基因控制,属温敏核不育。据此可以推断长S的育性受同一对隐性核基因控制,也属温敏核不育。
3 讨论
依据本文研究结果可知,长S虽来自普通野生稻与栽培稻的杂交后代,但其育性基因与C815S、广占63S、株1S、湘陵628S、HD9802S的育性基因等位,受1对隐性核基因控制,而与HN5S、培矮64S的育性基因不等位。C815S来自5SH038和培矮64S的杂交后代,而5SH038来自安农S-1的衍生后代,因而C815S可能具有安农S-1的不育基因或培矮64S不育基因,或者同时具有两者的不育基因[9-12]。安农S-1属温敏核不育,其不育性状受1对隐性核基因控制[8]。依据本文研究结果结合上述分析,可以推测C815S中与长S等位的不育基因应为来自安农S-1的不育基因。广占63S来自农垦58S的衍生后代,王宝和等人[14]研究表明广占63S的不育性状受1对隐性核基因控制,其育性转换主要表现为温敏型,与安农S-1的育性转换敏感性相似,但其育性基因与农垦58S是否等位尚未明确。由本文研究结果可推知广占63S的育性基因与农垦58S不等位,而与长S、安农S-1、株1S、HD9802S等温敏核不育系的育性基因等位,属温敏核不育。湘陵628S来自株1S,而株1S属温敏核不育,不育性状受1对隐性核基因控制[15-16],由本文研究结果可推知长S、株1S、湘陵628S的育性基因等位。周勇等[17]研究表明HD9802S属温敏核不育,不育性状受1对隐性核基因控制,且不育基因与温敏核不育系香125S(衍生自安农S -1)、株1S的育性基因等位,而与光温敏核不育系培矮64S和1103S(均衍生自农垦58S)的育性基因不等位。结合本文研究结果,可以推论长S育性基因与安农S-1的育性基因等位。综合上述结果,可以推论长S、广占63S、安农S-1、株1S、HD9802S虽来源不同,但育性均由同一对隐性核基因控制,均属温敏核不育。
陈立云等[19]认为温敏不育系的不育起点温度基本上是稳定的,其育性不受光照长度的影响或影响极其微小,但却受日平均温度和低温持续时间的影响;温敏不育系的育性要发生变化,必须要达到一定的低温积累量,积累量越大,育性变化的程度越明显;不同温敏不育系之间对低温持续时间的反应差异很大。结合本文研究,长S经历了8月上旬连续8天的低温后,育性没有发生变化,而经历了8月下旬的持续低温后,育性发生了明显变化,这说明长S的育性不仅受低温程度的影响,还受低温持续时间的影响。只有当低温达到一定程度并持续一定时间,长S的育性才会发生变化,这与陈立云等人的上述观点是一致的。这一结果还似乎说明能引起长S育性变化的低温程度并不是固定不变的,而与低温持续的时间之间存在一定的互补性。当低温持续时间较长时,即使低温程度偏高,也能引起长S的育性变化;反之,当低温持续时间较短时,即使低温程度偏低,长S的育性也难以发生变化。
由本研究结果可知,长S与目前生产上大面积利用的温敏不育系广占63S、株1S等育性基因等位。目前生产上这类温敏不育系的广泛利用,存在着极大的制种和繁殖风险。如2009年7月下旬的持续低温使长江中下游地区两系杂交稻制种大面积失败[20]。虽部分温敏核不育系的不育起点温度较低,降低了制种风险,但仍存在较大的繁殖风险。如温敏不育系2007年在海南繁殖时遇异常低温,2008年和2009年又遇异常高温,连续三年繁种失败[21]。本文研究结果表明,在长日条件下,长S需较长时间的持续低温育性才会发生变化,说明长S具有制种安全性,但长S可能存在较大的繁殖风险,因而需探索新的技术路线方能在生产上得以利用。李新奇等[22]提出将光敏不育基因与温敏不育基因重组,选育安全型温敏不育系的带有光敏不育基因的近等基因系,用其作为父本与安全型温敏不育系杂交,其F1代替安全型温敏不育系用于生产。这一技术路线可以较好地解决温敏不育系存在的制种和繁殖风险问题,为长S的利用提供了新思路。而陈立云等[23]认为在水稻两用核不育系中,不存在光敏不育基因和温敏不育基因,只存在1个或2个主效不育基因,其育性转换是主效不育基因与发育感光基因或(和)发育感温基因相互作用的结果。将温敏核不育系与强感光品种杂交,可以选育到不育起点温度低、光温互补作用强的光温敏核不育系,从而达到不育系制种安全且易繁殖的目的。这为长S的利用提供了另一种思路。
[1] 石明松. 晚粳自然两用系选育及应用初报[J].湖北农业科学,1981(7):1-3.
[2] 曹新国,胡锋,王美琴,等. 优质香型超高产两系中籼组合两优6326特征特性及应用[J].农艺科学,2008,24(9): 222-225.
[3] 斯华敏,刘文真,付亚萍,等. 我国两系杂交水稻发展的现状和建议[J].中国水稻科学,2011,25(5): 544-552.
[4] 郑卓,段世华,贺根和,等.水稻新资源光(温)敏核不育系长S的初步研究[J].贵州农业科学,2010,38(12): 1-3.
[5] 马来运,戴绍钧.湖农5S的选育及育性转换特性的研究[J].杂交水稻,1996,(1):3-5.
[6] 景润春.籼型光敏核不育水稻HN5S育性遗传研究[D].武汉:华中农业大学,1997.
[7] 张再君,梁承邺,戴绍钧.水稻光敏核不育系HN5S不育性的遗传分析[J].作物学报,2002,28(1):131-135.
[8] 邓华凤.舒福北.袁定阳.安农S-1的研究及其利用概况[J].杂交水稻,1999,14(3):1-3.
[9] 郭名奇.水稻两用不育系安湘S的选育及其应用初报[J].湖南农业科学,1995,(3):11-12.
[10] 郭武强,谭军,郭国强,等.籼型水稻温敏核不育系安湘S研究与应用概况[J].农业生物技术科学,2008,24(10): 106-109.
[11] 罗孝和,邱趾忠,李任华.导致不育临界温度低的两用不育系培矮64S[J].杂交水稻,1992,7(1):27-29.
[12] 唐文邦,陈立云,肖应辉,等.水稻两用核不育系C815S的选育与利用[J].湖南农业大学学报:自然科学版, 2007,33(8):26-31.
[13] 杨振玉,张国良,张从合,等.中籼型优质光温敏核不育系广占63S的选育[J].杂交水稻,2002,17(4):4-6.
[14] 王宝和,徐建军,吴银慧,等.水稻光温敏雄性核不育系广占63S不育基因的遗传分析与分子定位[J].中国水稻科学,2010,24(4):429-432.
[15] 杨远柱,符辰建,胡小淳,等.株1S温敏核不育基因的发现及超级杂交早稻育种研究[J].中国稻米,2007,(6): 17-22.
[16] 符辰建,秦鹏,胡小淳,等.水稻温敏核不育系湘陵628S的选育[J].中国农业科技导报,2010,12(6):90-97.
[17] 周勇,居超明,徐国成,等.优质早籼型水稻温敏核不育系HD9802S的选育与应用[J].杂交水稻,2008,23(2):7-10.
[18] 卢兴桂.中国光、温敏雄性不育水稻育性生态[M].北京:科学出版社,2002:135.
[19] 陈立云.两系法杂交水稻的理论与技术[M].上海:上海科学技术出版社,2001:102-105.
[20] 陆作楣.我国杂交稻育种研究的亮点及难点[J].中国水稻科学,2011,25(3):231-235.
[21] 陈立云,雷东阳,唐文邦,等.两系法杂交水稻研究和应用中若干问题的思与行[J].中国水稻科学,2010,24(6): 641-646.
[22] 李新奇,袁隆平,邓华凤,等.水稻光敏与温敏核不育基因之间互作效应与利用研究[J].科技导报, 2009,27(3): 74-79.
[23] 陈立云,肖应辉.水稻光温敏核不育机理设想及光温敏核不育系选育策略[J].中国水稻科学,2010,24(2): 103-107.
Alleism Test of Rice Male Sterile Gene in a New Thermosensitive Genic Male Sterile(TGMS) Line Chang S
*ZHENG Zhuo1,ZHANG Bei-ling,WANG Ai-ping1,WU Yang1,HE Li1,DUAN Shi-hua1,XU Dong-feng1,ZHENG Zi-wei1
(1.School of Life Science,Jinggangshan University, Ji’an, Jiangxi 343009, China;2. Ji’an City Agricultural Bureau, Ji’an, Jiangxi 343000, China)
Chang S is a new rice thermosensitive genic male sterile (TGMS) line obtained from the offsprings of the hybrid betweenand Zhenzhu’ai (rice). The fertility of Chang S is sterile under longer light duration with high temperature but fertile under lower temperature in field, and the conversion of the fertility is obvious. Fertile gene is allelic between Chang S and C815S, Guangzhan 63S, Zhu1S, Xiangling 628S, HD9802S, but is nonallelic between Chang S and HN5S, Pe′ai64S.
rice (L.); thermosensitive genic male sterility (TGMS); Chang S; allelism test
Q321+.8
A
10.3969/j.issn.1674-8085.2012.02.025
1674-8085(2012)02-0099-05
2011-11-11;
2012-02-12
江西省生态环境与资源重点实验室专项基金项目 (GJJ09658)
*郑 卓(1973-),男,湖北竹溪人,副教授,博士,主要从事水稻遗传育种研究(E-mail: zhengzhuodai@126.com);
张蓓玲(1969-),女,江西万安人,农艺师,主要从事农学研究(E-mail: jahxb@126.com);
王安萍(1971-),女,江西吉安人,实验师,硕士,主要从事遗传学研究(E-mail: jgkwangap@hotmail.com);
吴 杨(1980-),男,江西吉安人,讲师,博士,主要从事蔬菜逆境生理与分子生物学研究(E-mail: wuyangfenghao@126.com);
贺 俐(1981-),女,江西吉安人,实验师,硕士,主要从事蔬菜逆境生理与分子生物学研究(E-mail: heli@jgsu.edu.cn);
段世华(1968-),男,江西永新人,教授,博士,主要从事水稻分子生物学研究(E-mail: shihua_duan@yahoo.com.cn);
许东风(1965-),女,江西吉安人,教授,主要从事生物课程与教学论研究(E-mail: xudongfeng@jgsu.edu.cn);
郑自伟(1987-),男,湖北竹溪人,主要从事水稻遗传育种研究(E-mail: 375058635@qq.com).
