尼罗罗非鱼早期发育形态及其在珠江水系的空间分布
2012-10-12谭细畅李新辉李跃飞
谭细畅,李新辉,李跃飞,李 捷
中国水产科学研究院珠江水产研究所,广东广州510380
罗非鱼(tilapia),俗称非洲鲫鱼,隶属于鲈形目罗非鱼属,为广盐性鱼类。罗非鱼作为联合国粮农组织(FAO)向全世界推广养殖的优良品种之一,有多个品系,其养殖范围已遍布85个国家和地区。1956年我国从越南引进莫桑比克罗非鱼,经过养殖及推广,其在我国淡水养殖产量逐年增多,出口量也快速增长;2006年产量达到100万t,出口量超过18万t,出口额达3.7亿美元(朱华平等,2008)。然而,罗非鱼作为一种外来物种,在给我国带来良好经济价值的同时,对土著种类及水环境造成了较大影响。
目前,罗非鱼生物入侵的问题在国际上受到越来越多的关注。在美国、墨西哥以及巴西等国家和地区,罗非鱼已经成功入侵到淡水水体中(Kolar&Lodge,2002;Peterson et al.,2004)。卢旺达共和国Luhondo湖1935年引进罗非鱼,到1952年当地原有的3个鲤科种类基本消失(De Vos et al.,1999);瑙鲁共和国于1960年引进罗非鱼,到1979年罗非鱼便成为优势种类,原有的虱目鱼Chanos chanos Forsskal消失(Fortes,2005);尼加拉瓜1959年引进罗非鱼,1995年当地的细小银汉鱼Atherinella sardina Meek 基本消失(McCrary et al.,2007)。许多国家意识到引进的罗非鱼扩散到自然水体中对当地水生生物多样性的威胁,努力采取措施,防治及清除罗非鱼。然而,在我国,罗非鱼引入后的危害性很少受到关注,有关其早期发育过程的观察也仅是以生产及选育为目的,而忽略了其竞争性的生物学特征(王令玲和仇潜如,1981)。同时,罗非鱼从养殖水体逃逸后,在珠江水系扩散并建群,使珠江水系水生生物安全面临严峻威胁,但有关其具体扩散状况及其生态安全防范措施尚无报道。
笔者在近几年珠江水系鱼类资源调查中,经常采集到尼罗罗非鱼(以下简称罗非鱼)。本文通过对罗非鱼早期发育形态及其在珠江水系的空间分布进行研究,分析其入侵现状,为保护珠江水系鱼类生态及资源提供依据。
1 材料与方法
1.1 罗非鱼胚后发育观察
将野外采集到的罗非鱼成熟个体雌雄各3尾一同养殖在实验室玻璃箱(0.4 m ×0.6 m ×0.5 m)中,投喂普通颗粒饲料,水温23.0~28.8℃。养殖过程中成功获取其中1尾雌体口孵的鱼卵及鱼苗,达800尾(颗),观察该批鱼苗(卵)的早期发育形态。参照易伯鲁等(1988)对四大家鱼胚后发育时期的划分,对罗非鱼的10个主要发育期做详细的特征描述和拍照。测量各发育期鱼苗全长,计算其平均值。使用设备为XTZ-E型三目体视显微镜及尼康5400数码相机(上海光学仪器厂)。
1.2 罗非鱼苗的捕食性观察
将58.6~67.7 mm规格的罗非鱼苗养殖在玻璃缸(0.2 m ×0.3 m ×0.4 m)中,共 3 组,每组 15尾,水温26.0 ~28.5 ℃。投放20.0 ~26.1 mm 野外采集的鲢鱼Hypophthalmichthys molitrix Cuvier et Valenciennes苗,每天投喂1次,连续进行1周。统计罗非鱼苗的最大捕食量及捕食规格。
1.3 主要江段渔获物调查
西江调查江段为三水至柳江石龙,包括肇庆、梧州、藤县、桂平等地;北江调查江段主要为连江;东江调查惠州至河源江段,包括横沥、观音阁、古竹等地。调查方法为沿江随机抽查渔船,购买全船渔获物,记录时间、地点和捕捞渔具、网次,对渔获物进行种类鉴别,并测量其体长、体质量。重点调查三层流刺网的渔船,兼顾抛网、定置钩和虾笼等渔船。调查时间为2006~2010年的5~10月。
2 结果与分析
2.1 罗非鱼胚后发育形态
罗非鱼的卵呈浅黄色或金黄色,外形为鸭梨形,卵膜光滑透明(图1A、B)。受精卵吸水略为膨胀,其长径高2.77 mm、宽2.06 mm。
2.1.1 孵出期 出膜初期,全长5.7 mm,尾部动脉网丰富,卵黄囊上遍布血管(图1C)。卵黄囊比例很大,直径达2.24 mm,按照近似球体计算,体积约为3.3 mm3。养殖水温24℃下心脏搏动152~165 次·min-1。
2.1.2 眼黑期 出膜第4天,全长5.9 mm,出现明显口裂(图1D)。卵黄囊上仍遍布血管,但卵黄囊直径已缩小一半,仍呈球形。眼已黑,尾椎上翘,肠道有排泄物,出现雏形鳔,鳔上分布黑色素,头部背面、身体背部也分布黑色素。
2.1.3 背鳍分化期 全长6.1 mm,背鳍及臀鳍同时出现部分雏形鳍条,卵黄囊明显缩小,呈圆锥形,身体黑色素增多(图1E)。大部分罗非鱼贴鱼缸底部游动,少量可以向上游动到水面。
2.1.4 鳔一室期 全长7.4 mm,背鳍分化向前延伸,第二背鳍出现,卵黄囊进一步缩小,呈三角形(图1F、G)。肌节数13+18对,身体上分布许多黑色素,以头部背面、身体背部居多。
2.1.5 卵黄吸尽期 全长8.4 mm,卵黄囊完全消失,背鳍鳍条上出现明显大块黑色素斑(图1H)。腹鳍出现。
2.1.6 臀鳍形成期 全长12.5 mm,臀鳍已经形成(图1I)。
2.1.7 仔稚鱼期 全长13.6 mm,体形上已经接近成鱼(图1J),可以摄食其他种类的鱼苗,每尾每天可以摄食5~7尾鱼苗。
2.2 罗非鱼苗的捕食性
罗非鱼苗呈现很强的攻击性与捕食性。全长58.6~67.7 mm的罗非鱼苗单尾对鲢鱼苗的最大捕食量为12 ~20 尾·d-1;同时,观察发现,全长58.6 mm的罗非鱼苗未摄食全长26.1 mm的鲢鱼苗,初步认为其最大捕食规格为其全长的44.5%。

图1 尼罗罗非鱼早期发育形态图谱Fig.1 Morphology of small developmental stages tilapia in the Pearl River
2.3 珠江水系罗非鱼调查结果
在西江三水至柳江石龙所有调查江段均发现罗非鱼。在石龙江段刺网渔获物中的数量比为0.60%;在梧州江段刺网渔获物中的数量比为0.15%;在肇庆江段刺网渔获物中的数量比为1.00%。2010年10月3~5日在桂林兴安县灵渠三层流刺网的渔获物中,罗非鱼的数量比为4.00% ,质量比为 14.80% 。
罗非鱼在北江分支连江刺网、地笼及钓钩等网具渔获物中的数量比为0.95%。
罗非鱼在东江观音阁江段刺网渔获物中的数量比为 26.6%;体长为 13.0 ~ 28.5 cm,平均体长16.9 cm;体质量为 85 ~781 g,平均体质量 188.7 g。在惠州及河源江段渔获物中的数量比低于0.1%。
参考近年珠江水系鱼类资源调查结果(李捷等,2009;李跃飞等,2008;卢立,2005;韦日峰等,2009)可知,罗非鱼在珠江水系已经蔓延,局部江段为优势种(图2)。
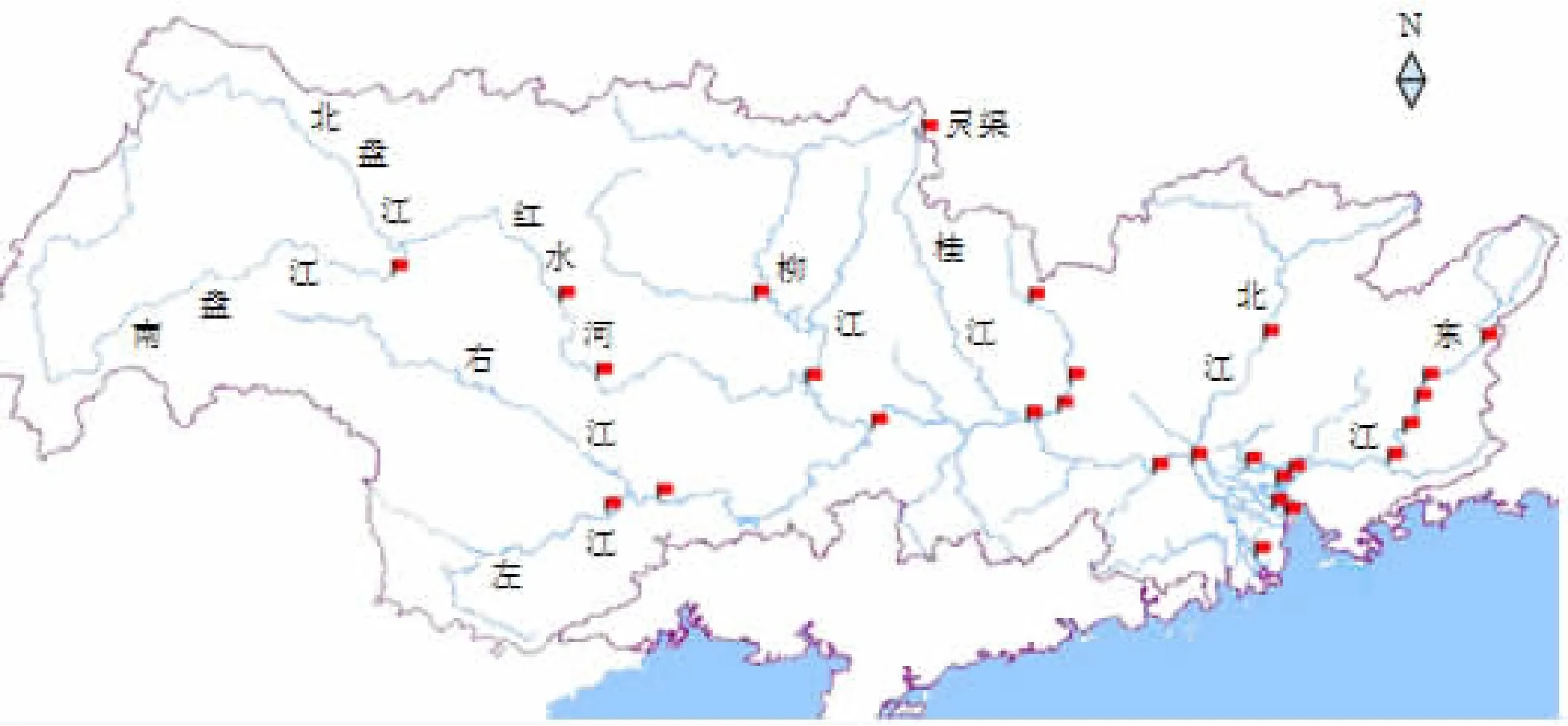
图2 罗非鱼在珠江水系的分布(旗帜符号标明分布)Fig.2 Distribution of tilapia in the Pearl River tributary(Flags mark the locations where tilapia was collected)
3 讨论
3.1 罗非鱼苗早期发育特征
罗非鱼苗早期发育特征:血液循环系统完善,血管网丰富;心脏搏动152~165次·min-1,比鲤Cyprinus carpio L.(120 次·min-1)(谭细畅等,2009)、厚颌鲂Megalobrama pellegrini Tchang(110次·min-1)(王剑伟等,2005)、鳜 Siniperca chuatsi Basilesky(150次·min-1)(郑闽泉等,1994)高;具有良好的适应性,氧气需求量小。卵黄囊体积大,类似于中华鲟Acipenser sinensis Gray,血管网密布;早期营养充足,内源营养期与外源营养期不需要过渡期,发育速度快,出膜第7天各鳍基本形成。
由于罗非鱼苗卵黄的营养远比四大家鱼及广东鲂Megalobrama terminalis Richardson的鱼苗丰富,且罗非鱼亲鱼口孵化鱼苗,并在出膜后有一定时期的护幼行为,使得罗非鱼苗有更强的适应性及竞争力。其优越的生物习性及入侵特征,比珠江水系土著种类更有竞争性,呈现很强的攻击性与捕食性。因此,应呼吁整个社会重视罗非鱼生物入侵问题,切实保护好珠江水系的鱼类资源及水生生态环境。
3.2 罗非鱼在珠江水系的分布
罗非鱼目前在珠江水系各支流西江、北江、东江都有分布,入侵情况较为严峻。在西江上游南盘江有分布(卢立,2005);在红水河龙滩水电站库区有分布(施军,2007);在左江的百色、阳纡有分布(周解,2002);在柳江及红水河中游有分布(韦日峰等,2009);在漓江暂未发现罗非鱼(蔡德所等,2009;朱瑜等,2007)。在东江的新丰江水库、枫树坝水库等也有分布(赵会宏等,2007)。在湖泊,如肇庆星湖、惠州市西湖、广州市公园湖泊已经成为优势种类,为重点清除对象。
灵渠位于珠江、长江两大水系的连接点,且在珠江水系的最北,该地有罗非鱼的分布。这说明一方面,温度已经不能限制罗非鱼在整个珠江水系的扩散;另一方面,长江水系也面临罗非鱼入侵的风险。目前,我国已进行耐寒罗非鱼品系的研究,一旦耐寒罗非鱼分布到长江,长江的鱼类群落将面临严峻风险。
根据2007~2009年广州市水生动植物本底调查,罗非鱼已经在除伶仃洋以外的各个水域建立了庞大的野生种群(赵峻等,2010)。在东江古竹江段,罗非鱼已经成为渔业捕捞的最主要种类;另外,根据该江段鱼类产卵场的调查,罗非鱼在鱼苗中的比例也高达9.7%(谭细畅等,2012)。因此,罗非鱼已经呈现泛滥的趋势。
为保护珠江水系原有的鱼类种质资源,有必要对珠江水系罗非鱼的分布进行更深入的调查,并监测各大支流的自然繁殖状况。在目前没有罗非鱼的分布江段,应该规划罗非鱼屏蔽区,禁止在该江段及邻近水域养殖罗非鱼,以防止其进入。另外,需要调查罗非鱼在一些江段泛滥的原因,进而采取有效措施控制其种群的进一步扩张。
致谢:感谢刘超及王培欣在试验中给予的帮助!
蔡德所,赵湘桂,朱瑜,周解,施军,韩耀全.2009.漓江鱼类资源调查及物种多样性分析.广西师范大学学报:自然科学版,27(2):130-136.
李捷,李新辉,谭细畅,李跃飞,何美峰,罗建仁,林建志,苏少芳.2009.广东肇庆西江珍稀鱼类省级自然保护区鱼类多样性.湖泊科学,21(4):556-562.
李跃飞,李新辉,谭细畅,李捷,王超,罗建仁.2008.西江肇庆江段渔业资源现状及其变化.水利渔业,28(2):80-83.
卢立.2005.广西金钟山自然保护区鱼类资源考察报告.湖南环境生物职业技术学院学报,11(4):319-324.
施军.2007.龙滩水电站库区鱼类资源调查研究报告.广西水产科技,(4):7-14.
谭细畅,李新辉,林建志,李跃飞,毕晔,李捷,王超.2009.珠江肇庆江段鲤早期发育形态及其补充群体状况.大连水产学院学报,24(2):125-129.
谭细畅,李跃飞,李新辉,李捷,王超.2012.梯级水坝胁迫下东江鱼类产卵场现状分析.湖泊科学,24(3):443-449.
王剑伟,谭德清,李文静.2005.厚颌鲂人工繁殖初报及胚胎发育观察.水生生物学报,29(2):130-136.
王令玲,仇潜如.1981.尼罗罗非鱼胚胎及胚后发育的观察.动物学报,27(4):327-336.
韦日峰,郑兰平,陈小勇,杨君兴.2009.广西河池地区鱼类资源调查及两支流的鱼类多样性比较.动物学研究,30(2):185-194.
易伯鲁,梁秩燊,余志堂.1988.葛洲坝水利枢纽与长江四大家鱼.武汉:湖北科技出版社.
赵会宏,崔科,甘炼,刘丽,孙际佳,李桂峰,王正印,梁浩亮,杨熙凯.2007.东江鱼类资源调查结果初报.华南农业大学学报,28(2):52-56.
赵俊,易祖盛,周先叶,肖智.2010.广州市水生动植物本底资源.北京:科学出版社.
郑闽泉,丁桂枝,黄涵生,刘伯仁,陈进荣.1994.鳜鱼胚胎发育的观察.江西水产科技,(3):21-25.
周解.2002.右江金鸡滩电站对渔业资源的影响和对策.广西水产科技,(3):1-15.
朱华平,卢迈新,黄樟翰.2008.罗非鱼健康养殖实用新技术.北京:海洋出版社.
朱瑜,周解,施军.2007.漓江的鱼类调查.广西水产科技,(2):66-78.
De Vos L,Snoeks J and van den Audenaerde D T.1999.The effects of tilapia introductions in Lake Luhondo,Rwanda.Environmental Biology of Fishes,27:303 -308.
Fortes R D.2005.Review of Techniques and Practices in Controlling Tilapia Populations and Identification of Methods That May Have Practical Applications in Nauru Including A National Tilapia Plan.New Caledonia:Secretariat of the Pacific Community Noumea.
Kolar C S and Lodge D M.2002.Ecological predictions and risk assessment for alien fishes in North America.Science,298:1233-1236.
McCrary J K,Murphy B R,Stauffer J R and Hendrix S S.2007.Tilapia(Teleostei:Cichlidae)status in Nicaraguan natural waters.Environmental Biology of Fishes,78:107 -114.
Peterson M S,Slack W T,Brown-Peterson N J and McDonald J L.2004.Reproduction in nonnative environments:establishment of nile tilapia,Oreochromis niloticus,in Coastal Mississippi Watersheds.Copeia,4:842 -849.
