广东省福寿螺形态性状变异及其与体质量的关系
2012-10-12李玺洋顾党恩牟希东罗建仁章家恩胡隐昌中国水产科学研究院珠江水产研究所农业部热带亚热带水产资源利用与养殖重点实验室广东广州50380华南农业大学热带亚热带生态研究所广东广州5064
李玺洋,徐 猛,罗 渡,顾党恩,牟希东,罗建仁,章家恩,胡隐昌*中国水产科学研究院珠江水产研究所,农业部热带亚热带水产资源利用与养殖重点实验室,广东广州50380;华南农业大学热带亚热带生态研究所,广东广州5064
福寿螺Pomacea canaliculata Lamarck原产于南美洲亚马逊河流域,由于其生长快、繁殖力强、易饲养、产量高等特点,在东南亚国家泛滥成灾,严重威胁水生农作物以及当地的生物多样性,已被列为入侵我国的16种恶性外来入侵物种之一。
1981年福寿螺被首次引入我国广东养殖,由于广东地区适宜其生长,且其适应性和耐受性很强(刘建康,1999),导致其大量繁殖,对广东的农业生产以及生态环境造成了严重的破坏。福寿螺变异快也是其能在入侵地快速扩张的原因之一(Alejandra et al.,2006),其形态随着栖息地的生长环境、生长区域密度、食物来源等不同有较大变异(李小慧等,2009;宋鄂平等,2006)。董胜张和俞晓平(2011)研究发现,食物种类可显著影响福寿螺的生长发育;叶建人等(2011)研究发现,取食率与福寿螺体质量增长呈显著正相关。关于福寿螺形态性状与体质量的关系也有部分报道(杨代勤等,1993;尹尚武等,2000),但应用相关分析、通径分析对福寿螺形态性状变异及其与体质量关系的研究尚未见报道。本文通过测量广东省7个地区福寿螺的壳高、壳宽、口宽、层高,研究其与体质量之间的关系,旨在为研究福寿螺入侵后的适应性进化机制提供基础数据。
1 材料与方法
1.1 材料来源与数量
在广东7个地区(清远、阳江、番禺、肇庆、潮汕、惠州和中山),用网随机采集稻田、农田、沟渠里生长的福寿螺,放塑料桶中带回实验室,鉴别雌雄(雌成螺的厣中央平滑略凹陷,雄成螺的厣中央外凸,且边缘向内微凹)并计算性比,采样数量和时间见表1。用游标卡尺测量福寿螺形态学数据(壳高、壳宽、口宽、层高),并用电子天平测定体质量(精确到0.01 g)。测量标准:壳高是指螺壳轴线的最大长度;壳宽是指壳体的最大直径;口宽是指壳口最大直径;层高是指壳体中最大一层壳层轴线的最大长度。
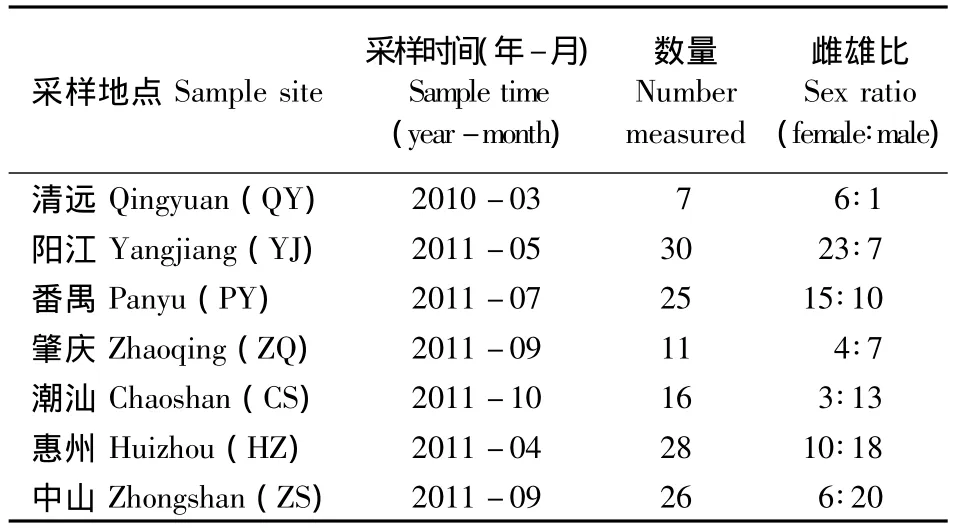
表1 不同采样点福寿螺的取样数量及性别比例Table 1 Number and sex ratio of apple snail in different sample sites in Guangdong Province,Southern China
1.2 数据分析
为使福寿螺形态数据与体质量满足正态分布,具有统计学意义,本文以LN函数对原始数据进行转换,然后进行统计学分析。运用统计软件Excel对所得数据进行分析,计算均值和标准差;以形态参数为自变量、体质量为依变量,应用统计软件SPSS 17.0分别进行相关分析、通径分析,计算出各自的相关系数、通径系数和决定系数。运用SPSS模板描述(describe)功能计算表型参数,应用模板相关(correlate)进行相关分析。通径系数中的直接系数,间接系数为∑rp,单独决定系数xixjjdyxi=P2yxi,共同决定系数dyxiyi=2rxiyiPyxiPyxi。式中,byx为自变量与依变量的偏回归系数;δx为自变量标准差,即形态数据标准差;δy为依变量标准差,即体质量标准差;rxy为person系数(顾万春,2006;李宁,2003)。
2 结果与分析
2.1 各形态性状的表型参数
由表2可以看出,7个地区福寿螺的形态变异波动程度相似,标准差数值相差较小。偏度和峰度接近于0(表3),代表样本呈正态分布,具有统计学意义。偏度是衡量实数随机变量概率分布的不对称性。偏度为正,意味着绝大多数的值位于平均数左侧;偏度为负,代表绝大多数值位于平均数右侧。峰度是变量值的集中程度。峰度<0,表示数值分布比较分散;峰度>0,则表示数值分布比较集中。由表3可以看出,番禺的峰度都为正数,说明番禺的形态数据比较集中,螺的大小一致;肇庆、中山的峰度都为负数,说明肇庆、中山的形态数据比较分散,螺的大小不一致。
变异系数大,表示其遗传多样性广,可适应环境变化的能力强。由表4可以看出,7个地区福寿螺体质量的变异系数较形态大。清远、阳江的变异系数比其他地区大,说明这2个地区福寿螺的环境适应能力强。
2.2 各性状间的表型相关系数
7个地区福寿螺4个形态性状间及与体质量间的相关性均达到极显著水平(P<0.01)。由表5可以看出,肇庆、潮汕、中山、清远和番禺福寿螺4个形态性状与体质量间的相关性较高,相关系数均超过0.9。肇庆和潮汕福寿螺壳宽与体质量最相关;中山、清远、阳江和番禺福寿螺与体质量最相关的形态性状相同,都是壳高;仅惠州福寿螺的口宽与体质量最相关。其余相关性状中,壳高与口宽的相关性很高,清远、阳江、番禺、肇庆、潮汕、惠州和中山福寿螺壳高与口宽的相关系数分别为0.900、0.747、0.900、0.986、0.930、0.961、0.946;壳高与壳宽的相关性次之,番禺、肇庆、潮汕、中山福寿螺壳高与壳宽的相关系数依次为 0.896、0.943、0.892、0.924。
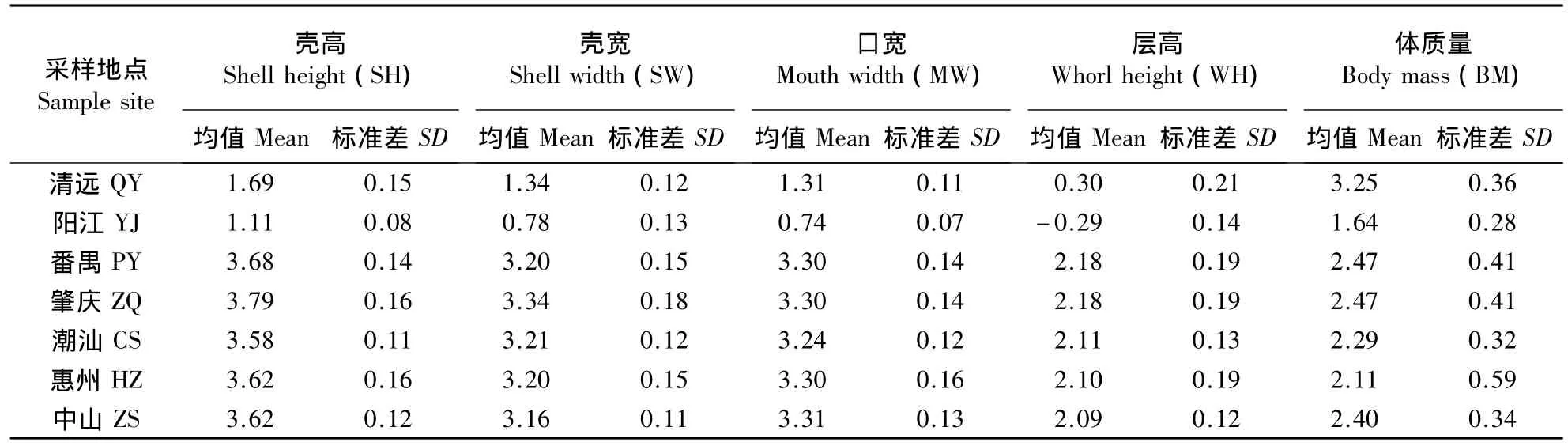
表2 广东省不同地区福寿螺形态参数的均值和标准差Table 2 Means and standard variations of morphological parameters of apple snail,P.canaliculata,at various locations in Guangdong Province,China

表3 广东省不同地区福寿螺形态参数的偏度和峰度Table 3 Skewness and kurtosis of morphological parameters of apple snail,P.canaliculata,at various locations in Guangdong Province,China
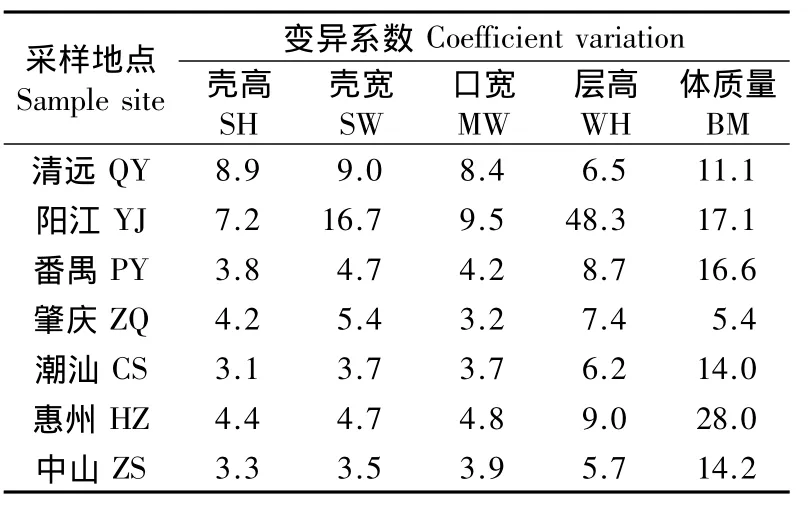
表4 广东省不同地区福寿螺形态数据的变异系数Table 4 Coefficient variation of morphological parameters of apple snail,P.canaliculata,at various locations in Guangdong Province,China

表5 广东省不同地区福寿螺形态性状与体质量的相关性Table 5 The relationship between four morphological traits and body mass for apple snail from various areas in Guangdong Province,Southern China
2.3 形态性状对体质量的通径系数
经过SPSS回归模型剔除P>0.05的参数后,发现7个地区福寿螺与体质量最相关的形态参数与用相关系数计算所得结果大致相同,即经过多元回归分析后,进一步确定仅壳高和壳宽与体质量最相关。通径分析建立在相关系数基础上进一步反映了原因与结果的真实关系。
清远、阳江地区福寿螺壳高对体质量的直接作用最大,远远大于另外几个形态性状对体质量的间接作用;番禺、中山虽然壳高与体质量最相关,但其直接作用小于间接作用;肇庆、潮汕壳宽与体质量最相关,且直接作用大于间接作用;惠州口宽与体质量最相关,且直接作用大于间接作用(表6)。
根据各性状对体质量的决定系数,计算出阳江壳高与壳宽对体质量的共同决定系数为0.1256,番禺壳高与层高的共同决定系数为0.2110,中山壳高与壳宽对体质量的共同决定系数为0.2435,清远壳高的单独决定系数为0.7534,肇庆壳宽的单独决定系数为0.4006,潮汕壳宽的单独决定系数为0.3831,惠州口宽的单独决定系数为0.5959。

表6 广东省不同地区福寿螺形态性状对体质量的影响Table 6 The effects of morphological traits on the body mass of the golden apple snail from 7 regions in Guangdong Province,China
3 讨论
5~10月福寿螺生长迅速,软体部分生长速度不及螺壳快,在螺壳停止生长后,软体部分还继续生长,直至填满整个螺壳。因此,螺壳越高,宽度越宽,其壳内空间就越大,软体部分有足够的空间生长,体质量也越大。杨代勤等(1993)通过测量508只福寿螺的壳高、壳宽和体质量,发现其壳高与壳宽呈直线相关,壳高与体质量、壳宽与体质量呈幂函数相关。本研究发现,除惠州之外,其余6个地区福寿螺的壳高、壳宽与体质量的相关性极高。因此,福寿螺的体质量与壳高呈正相关,测螺重,只需要测螺壳高即可(尹绍武等,2000)。
杨代勤等(1993)还指出,壳高、壳宽与层高之间呈直线关系,认为可以将层高作为测量福寿螺生长的一个较好的指标。由于福寿螺壳顶经常发生磨损,而层高随着寿命的延长而长高,测得的高度基本上能代表实际高度。因此,本研究中,番禺群体的层高与体质量较相关是可信的。
徐建荣(2009)研究了福建、广东、江苏的福寿螺,得出广东省福寿螺倾向于群体内相互交配,遗传多样性较低。本研究发现,清远的7只螺全部是黑色壳;阳江30只螺中只有3只是黑色,其余均是黄色;肇庆所采的11只螺全部是黄色。该结果也验证了徐建荣(2009)的结论。
本文仅是对福寿螺形态性状变异及其与体质量的相关性进行了探索性研究,未将这种变异与周围生长环境及食物来源进行深入分析。如果能系统地研究环境因素对福寿螺种群形态性状变异的影响,就可较为准确地根据形态性状推测其生长环境,并采取相应措施对其进行控制。因此,有关福寿螺生长发育方面的研究仍需深入。
董胜张,俞晓平.2011.食物种类与饥饿胁迫对福寿螺生长发育的影响.生物安全学报,20(3):213-219.
顾万春.2006.统计遗传学.北京:科学出版社.
李宁.2003.动物遗传学.北京:农业出版社.
李小慧,胡隐昌,宋红梅,王培欣,汪学杰,牟希东,刘超,罗建仁.2009.中国福寿螺的入侵现状及防治方法研究进展.中国农学通报,25(14):229-232.
刘建康.1999.高级水生生物学.北京:中国科学院.
宋鄂平,于吉涛,袁用道,胡胜华,曾克峰.2006.福寿螺入侵浙西南地区原因与防治方法.湖北农业科学,45(6):804-806.
徐建荣.2009.福寿螺繁殖生态与遗传多样性研究.青岛:中国海洋大学.
杨代勤,陈芳,刘百韬,李道霞.1993.福寿螺繁殖和生长的初步研究.湖北农学院学报,14(1):40-44.
叶建人,冯永斌,林贤文,王华弟,祝增荣.2011.福寿螺的寄主植物及其对福寿螺体重的影响.生物安全学报,20(2):124-131.
尹绍武,颜亨梅,王洪全,许芳.2000.福寿螺的生物学研究.湖南师范大学自然科学学报,23(2):76-82.
Alejandra L,Estebenet P R M and Silvana B.2006.Conchological variation in Pomacea canaliculata and other South A-merican Ampullariidae(Caenogastropoda,Architaenioglossa).Biocell,30:329 -335.
