典型草原三种蝗虫种群竞争关系的研究
2012-09-28吴惠惠徐云虎曹广春格希格都仁刘朝阳额尔登巴图王广君张泽华
吴惠惠, 徐云虎, 曹广春, 格希格都仁, 刘朝阳,贺 兵, 额尔登巴图, 王广君, 张泽华*
(1.中国农业科学院植物保护研究所,农业部锡林郭勒草原有害生物科学观测实验站,北京 100193;2.内蒙古自治区锡林郭勒盟镶黄旗草原工作站,锡林郭勒 013250)
蝗虫是草原生态系统中的初级消费者[1],其分布具有明显的集团性[2]。蝗虫本身的特性及竞争、栖境的特性、食物的有效性是决定蝗虫种群动态和分布的重要因素[3-4]。栖境结构的组成及变化,对蝗虫种群数量产生重要影响,同时,在适合蝗虫生存的栖境中,随着种群密度的增加,栖境质量将下降,则反过来影响蝗虫的栖境选择。尽管蝗虫种间存在着明显的食物分化[1,5],但蝗虫种群密度及其所利用食物的有效性和质量性的周期性波动,有可能提供竞争的机会,至少是间歇性的[6]。而蝗虫种群之间的竞争作用不仅可以通过降低出生率、增加死亡率的方式影响蝗虫种群数量动态[7],而且可以在个体水平上改变蝗虫生长的速度和型态,从而影响蝗虫种群的生活模式[8]。总之,蝗虫种内与种间竞争的关系模式,蝗虫栖境选择与种间竞争的关系是研究蝗虫种群数量动态变化的关键因子。
内蒙古锡林郭勒盟典型草原优势种亚洲小车蝗(Oedaleus asiaticus B.-Bienko)、毛足棒角蝗[Dasyhippus barbipes(F.-W.)]和鼓翅皱膝蝗[Angaracris barabensis(Pall.)]的生物学特征、生活习性已有大量研究[9-11]。所以本文在前人研究的基础上,比较分析了3种蝗虫的取食特性,同时采用野外罩笼方法,调查了不同密度下3种蝗虫成虫的死亡率,并设计正交试验分析不同密度及不同栖境下3种蝗虫的死亡率,旨在掌握典型草原3种蝗虫不同密度下群落结构和功能变化、种间竞争和栖境选择,预测群落的发展演替动态,同时也为草原蝗虫的有效防治提供科学依据。
1 材料与方法
1.1 试验区自然概况
试验区位于内蒙古自治区锡林郭勒盟镶黄旗草原,分布于北纬42°15′~42°25′,东经113°45′~113°83′。地势较为平坦,海拔高度相差无几。该地区属于中温带半干旱大陆性季风气候,年降水量267.9mm。地区年平均气温3.1℃,无霜期平均为120d左右。主要土壤类型为栗钙土。
试验选择3种草地类型,分别为Ⅰ:克氏针茅草原(Stipa krylovii-steppe):主要植被类型为克氏针茅(Stipa krylovii Roshev.),散生糙隐子草[Cleistogenes squarrosa(Trin.)Keng]、小叶锦鸡儿(Caragana microphylla Lam.)、银灰旋花(Convolvulus ammannii Desr.)、冷蒿(Artemisia frigida Willd.)等植物,在该样地进行亚洲小车蝗和鼓翅皱膝蝗的密度试验及正交试验;Ⅱ:羊草草原(Leymus chinensis-steppe):主 要 植 被 类 型 为 羊 草 [Leymus chinensis(Trin.)Tzvel.],散生糙隐子草、小叶锦鸡儿、银灰旋花、冷蒿、木地肤[Kochia prostrata(Linn.)Schrad.]、克氏针茅等植物,在该样地进行毛足棒角蝗的密度试验及正交试验;Ⅲ:克氏针茅富含杂类草草原(Steppe dominated by Stipa krylovii with the forb diverse):主要植被类型为克氏针茅、栉叶蒿[Neopallasia pectinata(Pall.)Poljak.]、二裂叶委陵菜(Potentilla bifurca Linn.)等,散生糙隐子草、小叶锦鸡儿、冷蒿等植物,在该样地进行正交试验。
1.2 蝗虫食性研究
选择克氏针茅、羊草、糙隐子草、小叶锦鸡儿、银灰旋花、冷蒿、栉叶蒿和木地肤作为供试牧草,保持新鲜,称取相同重量牧草,分别对3种蝗虫进行单独的连续强迫饲喂,观察蝗虫取食及死亡情况。
1.3 3种蝗虫单种群不同密度的田间死亡率
网捕亚洲小车蝗、毛足棒角蝗和鼓翅皱膝蝗成虫,按雌雄比1∶1放入罩笼 (1m×1m×1m),亚洲小车蝗和毛足棒角蝗的虫口密度分别为6、16头/m2和26头/m2,鼓翅皱膝蝗的虫口密度为4、6头/m2和10头/m2,3次重复。各罩笼内植被类型和最初生物量大致相同。试验从2011年6月25日开始,到7月15日结束,共20d,每5d检查1次,记录死亡数,并补充与死亡蝗虫相同数量的蝗虫。
1.4 3种蝗虫混合种群死亡率正交试验
采取4因素3水平正交试验,因素A为亚洲小车蝗虫口密度,水平为6、10头/m2和16头/m2;因素B为毛足棒角蝗虫口密度,水平为6、10头/m2和16头/m2;因素C为鼓翅皱膝蝗虫口密度,水平为4、6头/m2和10头/m2;因素D为不同的Ⅰ、Ⅱ、Ⅲ号样地。2011年6月25日网捕亚洲小车蝗、毛足棒角蝗和鼓翅皱膝蝗相应数目成虫,按雌雄比1∶1放入罩笼(1m×1m×1m),10d后记录存活数,计算死亡率。
1.5 数据处理
蝗虫种内密度试验采用多重比较和简单相关方法分析。
正交试验的试验数据采用极差分析和方差分析。
以上数据用SAS8.0软件统计分析。
2 结果
2.1 食性分析
亚洲小车蝗和毛足棒角蝗食性相似,嗜食羊草、糙隐子草和克氏针茅,不食小叶锦鸡儿、银灰旋花、木地肤和冷蒿;而鼓翅皱膝蝗食性杂,对禾本科牧草嗜好程度没有亚洲小车蝗和毛足棒角蝗强。
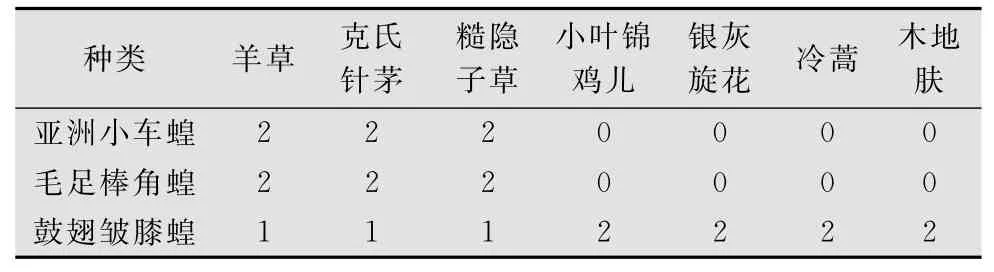
表1 3种草原蝗虫食性选择1)
2.2 3种蝗虫密度试验田间死亡率
多重比较结果表明,不同密度处理5d后,亚洲小车蝗、毛足棒角蝗和鼓翅皱膝蝗高、中、低密度间死亡率差异不显著,但是鼓翅皱膝蝗死亡率比其他两种蝗虫显著偏高(p<0.05);处理10d后,亚洲小车蝗和毛足棒角蝗的死亡率高密度(26头/m2)时比其中密度(16头/m2)和低密度(6头/m2)时显著增高(p<0.05),鼓翅皱膝蝗各密度之间的死亡率差异不显著;处理15d和20d后,亚洲小车蝗的死亡率低密度(6头/m2)时与其高密度(26头/m2)时相比显著降低(p<0.05),毛足棒角蝗和鼓翅皱膝蝗的死亡率各密度处理之间差异不显著(表2)。
亚洲小车蝗连续低密度(6头/m2)处理5、10、15、20d时,死亡率差异不显著;中密度(16头/m2)处理5、10d死亡率差异不显著,但是随时间延长,罩笼内植物减少,15、20d时死亡率显著增加(p<0.05);高密度(26头/m2)处理5d死亡率为零,10、15、20d死亡率显著增加(p<0.05),但彼此之间差异不显著。由表2可知Ⅰ号样地亚洲小车蝗的承载量约为11头/m2。
毛足棒角蝗连续低密度(6头/m2)处理5、10、15、20d时,死亡率差异不显著;中密度(16头/m2)和高密度(26头/m2)处理死亡率均随时间延长而显著增加(p<0.05),但15、20d时的死亡率差异不显著。由表2可知Ⅱ号样地毛足棒角蝗的承载量为16头/m2。
鼓翅皱膝蝗各密度处理试验开始时死亡率较高,均超过50%,但彼此之间差异不显著,并且鼓翅皱膝蝗死亡率与试验处理时间及密度相关性均不显著。

表2 罩笼内3种不同蝗虫的死亡率比较1)
2.3 混合种群正交试验
正交试验亚洲小车蝗死亡率结果表明,4个因素中,亚洲小车蝗虫口密度及样地环境对亚洲小车蝗死亡率的影响呈极显著水平(p<0.01),且样地环境对其影响比亚洲小车蝗虫口密度大;毛足棒角蝗虫口密度对亚洲小车蝗死亡率的影响呈显著水平(p<0.05);鼓翅皱膝蝗的虫口密度对亚洲小车蝗死亡率的影响不显著。亚洲小车蝗和毛足棒角蝗为低密度的Ⅱ号样地组合,亚洲小车蝗死亡率最低;亚洲小车蝗为中密度,毛足棒角蝗为高密度的Ⅰ号样地组合,亚洲小车蝗死亡率最高(见表3、表4)。
正交试验毛足棒角蝗死亡率结果表明,4个因素中,样地环境对毛足棒角蝗死亡率的影响呈极显著水平(p<0.01),其余3个因素对其影响不显著。毛足棒角蝗死亡率在Ⅱ号样地最低,Ⅲ号样地最高(见表5、表6)。
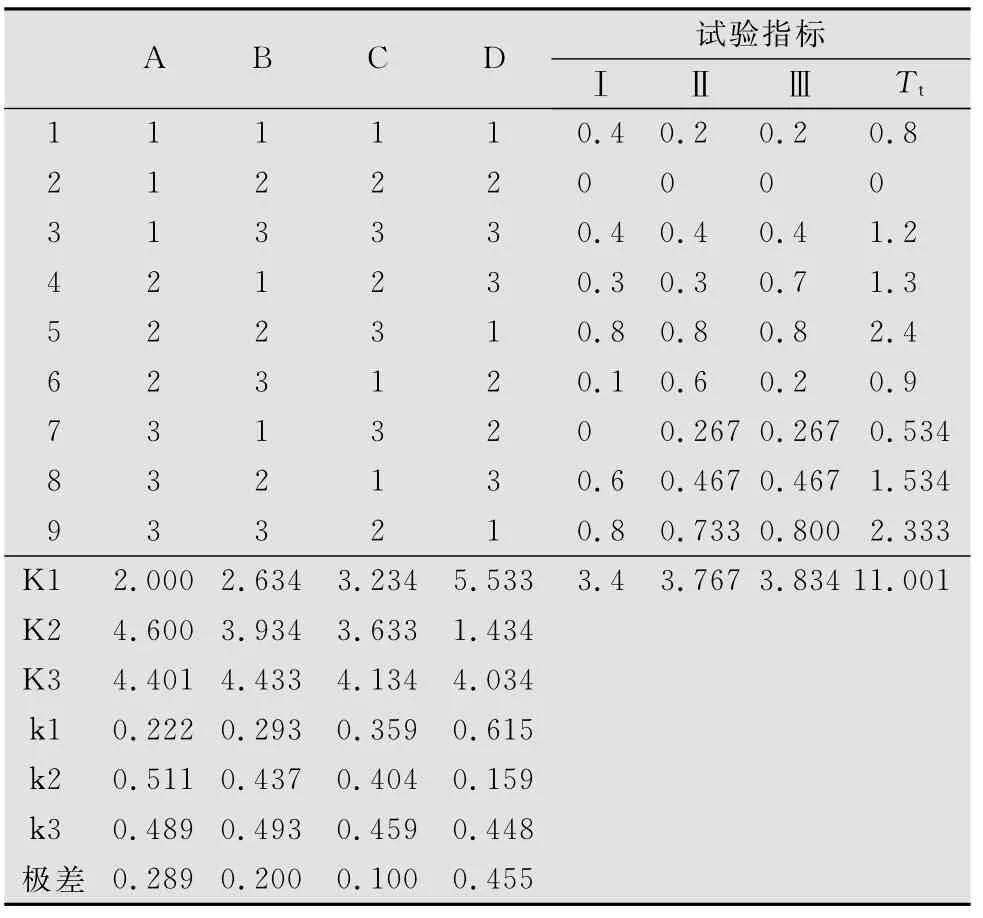
表3 亚洲小车蝗死亡率正交试验结果1)

表4 亚洲小车蝗死亡率正交试验方差分析1)
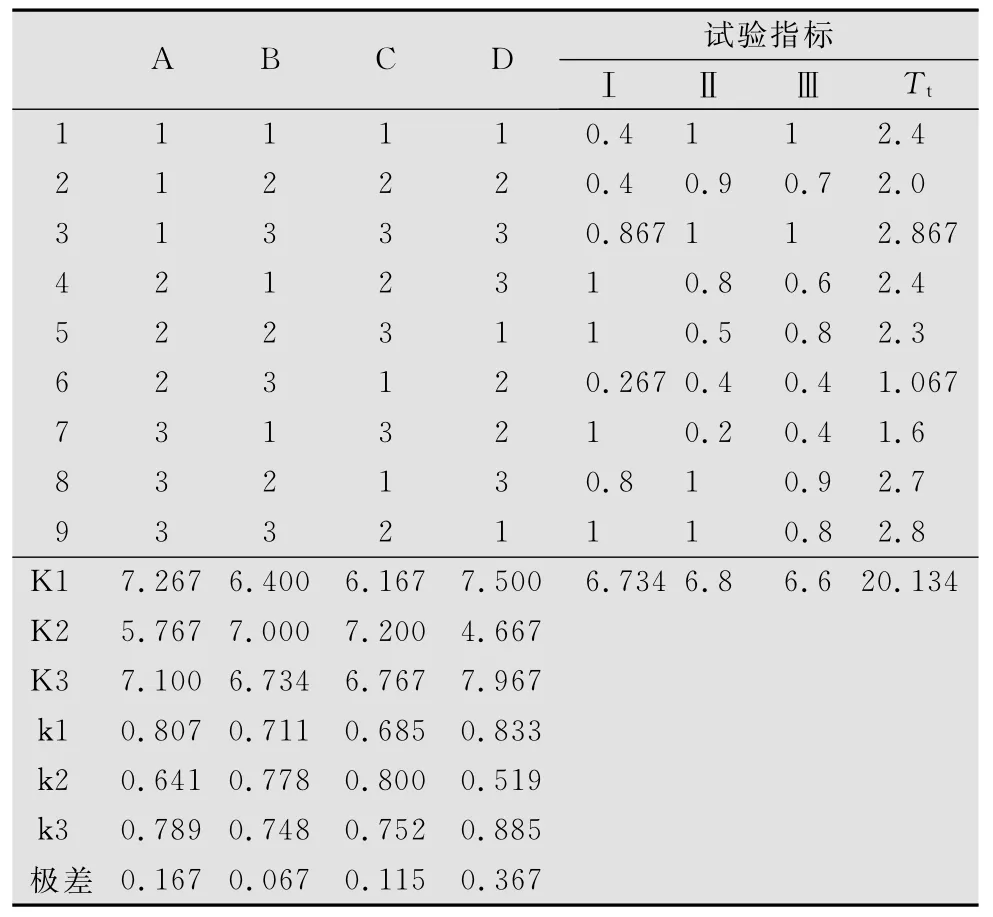
表5 毛足棒角蝗死亡率正交试验结果1)
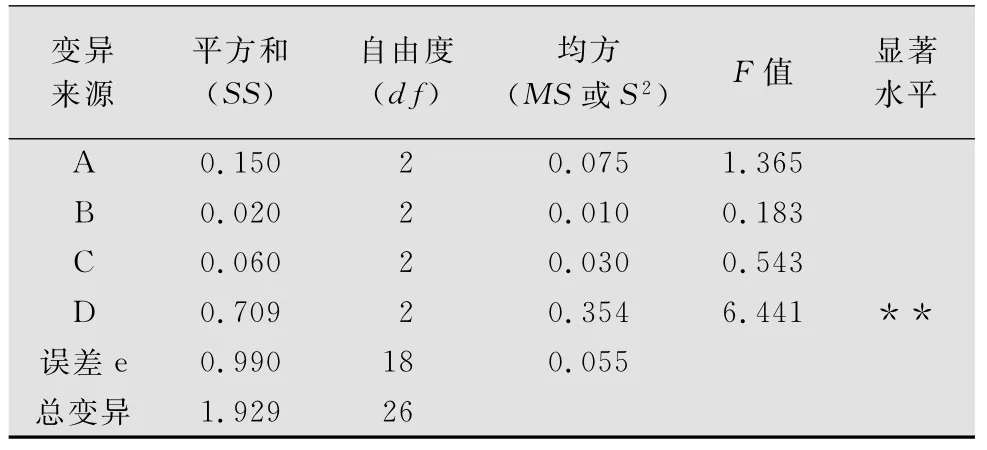
表6 毛足棒角蝗死亡率正交试验方差分析1)
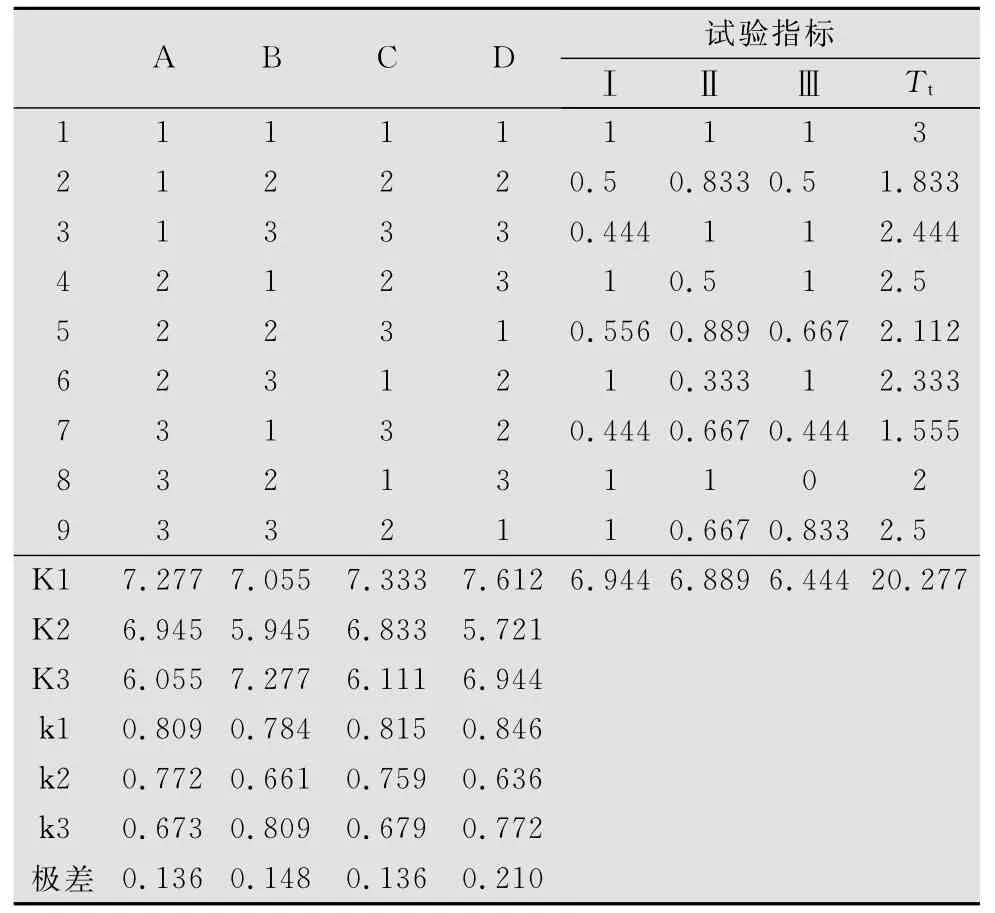
表7 鼓翅皱膝蝗死亡率正交试验结果1)
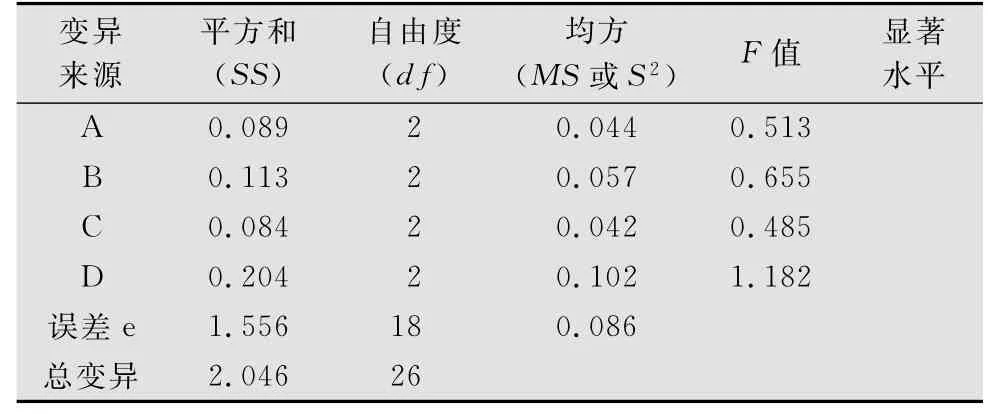
表8 鼓翅皱膝蝗死亡率正交试验方差分析1)
正交试验鼓翅皱膝蝗死亡率结果表明,4个因素中没有对鼓翅皱膝蝗死亡率的影响达显著水平的因素(见表7、表8)。
3 讨论
3.1 食物对种内竞争的影响
亚洲小车蝗和毛足棒角蝗食性相似,以禾本科牧草为主,而鼓翅皱膝蝗对于禾本科牧草偏嗜程度较差[11]。罩笼内前15d鼓翅皱膝蝗死亡率显著高于另外两种蝗虫 (p<0.05),而亚洲小车蝗和毛足棒角蝗之间的死亡率差异不显著。说明鼓翅皱膝蝗可利用的食物资源少,种内竞争激烈,因而死亡率急剧升高,而亚洲小车蝗和毛足棒角蝗在有充足的食物来源时,死亡率较低,可见食物限定显著影响蝗虫死亡率。
3.2 种群密度对竞争的影响
一般认为,在高密度的条件下可使蝗虫死亡率增高,所以通过密度依赖因素的作用,实现密度调节。围绕食物、空间的种内竞争是最基本的调节机制[12-13]。本试验中,高密度时,3种蝗虫种内竞争最强烈,亚洲小车蝗和毛足棒角蝗在试验的20d内死亡率一直呈上升趋势,最高分别达65.3%和41.3%,而鼓翅皱膝蝗平均死亡率一直居高不下,最高达85.2%;中密度时,各种群也存在较强的种内竞争,15d之后,各密度蝗虫种群持续为害,罩笼内食物减少,亚洲小车蝗和毛足棒角蝗的种内竞争增加,其死亡率较低密度下死亡率显著升高;低密度时,亚洲小车蝗和毛足棒角蝗种内竞争趋于平缓,种群密度下降缓慢。上述结论符合密度自我调节假说[14]。
在试验过程中,鼓翅皱膝蝗各密度处理死亡率一直较高,原因可能在于该样地并不是鼓翅皱膝蝗的理想样地,试验设计的4、6头/m2和10头/m2的密度水平对于鼓翅皱膝蝗均较高。鼓翅皱膝蝗与亚洲小车蝗、毛足棒角蝗所适宜的生境条件有所不同,需进一步对其栖境选择进行试验摸索。
3.3 栖境对蝗虫种群竞争的影响
混合种群正交试验10d后各蝗虫死亡率表明,亚洲小车蝗的种间竞争力最强(各处理平均死亡率为40.7%),而毛足棒角蝗和鼓翅皱膝蝗种间竞争力较弱(74.6%、75.1%)。混合种群中,亚洲小车蝗与毛足棒角蝗之间存在食物的竞争,但由于毛足棒角蝗个体较小,对栖境选择相对严格[3],在Ⅱ号样地之外的其他样地较难获得生存空间,与亚洲小车蝗的种间竞争表现较弱。但是在毛足棒角蝗最适合生存的Ⅱ号样地,其平均死亡率(51.9%)低于鼓翅皱膝蝗的平均死亡率(63.6%)。
正交试验结果表明,内蒙古锡林郭勒盟典型草原上,亚洲小车蝗是明显的优势种。亚洲小车蝗的栖息环境和自身虫口密度是影响其分布和种群动态的主要因素,与其存在食物竞争关系的毛足棒角蝗的虫口密度亦可影响其种群动态;毛足棒角蝗对栖境要求严格,在其适合的栖境之外死亡率增加,其自身虫口密度及其他种群的虫口密度也就无法对其死亡率构成显著性影响;而由于食物有效性和栖境特性的双重原因[15],混合种群中,鼓翅皱膝蝗降低密度以适应种间竞争。
[1] Kang L,Chen Y L.Multidimensional analysis of resource utilization in assemblages of rangeland grasshoppers(Orthoptera:Acrididae)[J].Entomologia Sinica,1994,1(3):264-282.
[2] He D H.The occurrence and advance of research on locusts and grasshoppers in the Northern grasslands of China[J].Entomological Research,1994,1:228-234.
[3] 颜忠诚,陈永林.草原蝗虫的栖境选择:栖境选择与水平结构的关系[J].武夷科学,1998(1):251-257.
[4] Belovsky G E,Slade J B.Dynamics of two Montana grasshopper populations:relationships among weather,food abundance and intraspecific competition[J].Oecologia,1995,101(3):383-396.
[5] 康乐,陈永林.草原蝗虫营养生态位的研究[J].昆虫学报,1994,37(2):178-189.
[6] Joern A,Klucas G.Intra-and interspecific competition in adults of two abundant grasshopper(Orthoptera:Acrididae)from a sandhills grassland[J].Environmental Entomology,1993,22(2):352-361.
[7] Belovsky G E.Optimal foraging and community structure:the allometry of herbivore food selection and competition[J].Evolutionary Ecology,1997,11(6):641-672.
[8] Beckerman A P.Counterintuitive outcomes of interspecific competition between two grasshopper species along a resource gradient[J].Ecology,2000,81(4):948-957.
[9] 李鸿昌,席瑞华,陈永林.内蒙古典型草原蝗虫食性的研究I笼罩供食下的取食特征[J].生态学报,1983,3(3):214-228.
[10] 颜忠诚,陈永林.内蒙古草原蝗虫个体大小及生活型划分的探讨[J].生态学报,1997,17(6):666-670.
[11] 卢辉,余鸣,张礼生,等.不同龄期及密度亚洲小车蝗取食对牧草产量的影响[J].植物保护,2005,31(4):55-58.
[12] 胡玲玲,刘勇,徐洪富,等.桃蚜、萝卜蚜的种内密度和种间竞争效应[J].华东昆虫学报,2004,13(1):77-80.
[13] 王健立,李洪刚,冯志国,等.西花蓟马与烟蓟马在紫甘蓝上的种间竞争[J].中国农业科学,2011,44(24):5006-5012.
[14] Wynne-Edwards K E.Biparental care in Djungarian but not Siberian dwarf hamsters(Phodopus)[J].Animal Behaviour,1995,50(6):1571-l585.
[15] 万秀莲,张卫同.草原毛虫幼虫的食性及其空间格局[J].草地学报,2005,13(1):84-88.
