致倦库蚊防御素基因的克隆与原核表达及蛋白纯化
2012-09-26王吉平张春林翟素珍
王 赟,王吉平,张春林*,翟素珍
(1.贵阳医学院医学生物技术教研室,贵州贵阳550004;2.贵阳医学院生物学教研室,贵州贵阳550004)
昆虫防御素(Defensin)术语最初用于Sarcophaga peregina和Phormia terranovae的两个抗革兰阳性菌抗菌肽,并因与哺乳动物噬菌细胞的一组抗菌肽序列相似而命名为防御素[1]。防御素作为天然免疫的效应分子为生物抵御病原菌的感染提供有效的第一位的防护。其中,昆虫防御素是昆虫受到意外伤害或微生物感染时,在血淋巴或消化道中产生的一类抗菌肽。昆虫防御素通过天然的免疫机制对免疫防御体系不完善的昆虫起着重要的作用[2]。
蚊虫在受到外来病原侵袭时,会激活其内源的防御素基因的表达,从而起到防御的作用[3]。致倦库蚊属于昆虫纲双翅目,不仅对人及牲畜骚扰吸血,而且是多种疾病的传播媒介,如日本脑炎、班氏丝虫病,其幼虫孳生于污染的水体,如粪坑、水坑、水沟等,携带病原体数量多,而自身却并不生病,因而推测其体内具有强大的防御系统。
本研究通过RT-PCR技术获得了致倦库蚊防御素基因全长编码区,构建含有致倦库蚊防御素成熟肽段的重组表达质粒p ET32a-DEF,并优化表达菌的培养、诱导条件,以实现重组蛋白的高效表达,最后获得了纯化的重组蛋白,这为进一步研究该蛋白的功能奠定了基础。
1 材料与方法
1.1 材料
1.1.1 试验用动物 致倦库蚊(贵阳株)于2010年采集于贵州贵阳,由贵阳医学院生物学教研室饲养繁殖。
1.1.2 菌种和质粒 原核表达载体p ET32a(+)及大肠埃希菌(Escherichia coli)Rosetta、DH5α由贵阳医学院生物学教研室保存,克隆载体p MD18-T购自宝生物工程(大连)有限公司。
1.1.3 主要试剂 DNA Marker、限制性内切酶Bam HⅠ和 Hin dⅢ、T4DNA连接酶、PCR TaqTM酶、蛋白质Marker为宝生物工程(大连)有限公司产品;总RNA提取试剂Trizol,质粒提取试剂盒为北京天根生物技术有限公司产品;第一链cDNA合成试剂盒为MBI公司产品;IPTG(isopropylthjo-β-D-galactoside,异丙基硫代-β-D-半乳糖苷)、氨苄青霉素、氯霉素为北京索莱宝科技有限公司产品;His-镍蛋白纯化套装为北京天恩泽基因科技有限公司产品;引物由上海生工生物工程技术服务有限公司合成。
1.2 方法
1.2.1 致倦库蚊防御素基因的获取 以冈比亚按蚊防御素氨基酸序列(ABB00947)为探针,对致倦库蚊EST序列库进行tBLASTn检索,得到1条与之同源的序列。将此序列用ORF Finder查找开放阅读框,并进行Blast比对为潜在防御素编码基因。
1.2.2 致倦库蚊防御素基因的扩增、克隆及鉴定根据比对获得的致倦库蚊防御素基因片段设计引物,上 游 Def F:5′-AGATGAACTCGCTTGGAA-3′,下游 Def R:5′-GGCCAAACAATATTTATTA-3′,由上海生工生物工程技术服务有限公司合成。采用Trizol法提取致倦库蚊总RNA,用紫外分光计测定总RNA纯度和浓度,cDNA第一链的合成按照Fermentas第一链合成试剂盒操作说明进行。目的片段扩增条件:94℃5 min;94℃1 min,50℃30 s,72℃1 min,30个循环;72℃5 min。扩增产物经琼脂糖凝胶电泳,回收目的片段,并与p MD-18T载体进行连接、转化大肠埃希菌DH5α;蓝白斑筛选阳性克隆并经菌落PCR验证后,送上海生工生物工程技术服务有限公司测序,鉴定正确的重组质粒命名为p MD-DEF。
1.2.3 致倦库蚊防御素成熟肽基因片段的PCR扩增 根据测序获得的防御素基因序列,用Primer Premier 5.0软件设计扩增成熟肽序列的上下游引物。上游引物CQF为:5′-CGGGATCCTTCCCTCAGGAGTC-3′,下划线为Bam HⅠ酶切位点;下游引 物CQR为:5′-CCCAAGCTTGTCAGTTTCGGCAGACGC-3′下划线为Hin dⅢ酶切位点。预计扩增长度为249 bp。以重组质粒p MD-DEF为模版,PCR扩增条件:94℃5 min;94℃1 min,60℃30 s,72℃1 min,30个循环;72℃5 min。PCR扩增产物经10 g/L琼脂糖凝胶电泳检测后,使用琼脂糖凝胶回收试剂盒回收目的片段。
1.2.4 重组表达质粒p ET32a-DEF的构建及测序分别用Bam HⅠ和Hin dⅢ双酶切p ET32a(+)质粒和防御素成熟肽基因片段。酶切产物经10 g/L琼脂糖凝胶电泳检测,用琼脂糖凝胶回收试剂盒回收酶切产物。用T4 DNA连接酶将质粒与目的基因片段连接,转化大肠埃希菌DH5α感受态细胞,挑取单菌落进行培养,经PCR初步筛选阳性克隆,再抽提重组质粒用Bam HⅠ和Hin dⅢ双酶切进一步鉴定,最后送上海生工生物工程技术服务有限公司进行测序。测序结果正确的命名为p ET32a-DEF。
1.2.5 重组表达质粒p ET32a-DEF在大肠埃希菌中的表达 分别将空载体p ET32a(+)和重组质粒p ET32a-DEF转化至大肠埃希菌Rosetta感受态细胞中,涂布于含氨苄青霉素(100μg/mL)、氯霉素(25μg/mL)的LB琼脂培养平板上,次日挑单菌落接种于含氨苄青霉素和氯霉素的LB液体培养基中,37℃振荡培养至OD600 nm约为0.6时,加入终浓度为1 mmol/L IPTG诱导4 h后,按常规进行SDS-PAGE检测,分析目的蛋白表达情况。
1.2.6 重组菌株诱导条件的优化方法 为了获得大量重组蛋白,以含100μg/mL氨苄青霉素、25μg/mL氯霉素的LB液体培养基作为培养和诱导表达的基本条件,对IPTG浓度、诱导时间和诱导温度进行了研究。
1.2.6.1 IPTG诱导浓度的优化 取过夜培养的含重组质粒的Rosetta菌,按1∶50稀释到含抗生素的LB液体培养基中,37℃,180 r/min振荡培养至OD600在0.6左右,分别加入终浓度为0、0.2、0.4、0.6、0.8、1.0、1.2、1.4 mmol/L 的IPTG,继续诱导4 h后取样,12%SDS-PAGE电泳分析重组蛋白的表达情况。
1.2.6.2 诱导时间的优化 取过夜培养的含重组质粒的Rosetta菌,按1∶50稀释到含抗生素的LB液体培养基中,37℃以180 r/min振荡培养至OD600 nm在0.6左右,以上一步确定的最佳IPTG浓度进行诱导,分别于诱导后0、1、2、3、4、5、6、7 h取样,12%SDS-PAGE电泳分析重组蛋白的表达情况。
1.2.6.3 不同诱导温度下重组蛋白表达形式的检测 取过夜培养的含重组质粒的Rosetta菌,按1∶50稀释到含抗生素的LB液体培养基中,37℃以180 r/min振荡培养至OD600在0.6左右,以前两步确定的最佳IPTG浓度、最佳诱导时间,分别于20、25、30、37℃进行诱导表达。诱导结束后,离心收集菌体,菌体沉淀重悬于细菌裂解缓冲液(50 mmol/L Tris-HCl p H 8.0、1 mmol/L EDTA、0.1 mol/L NaCl)中,反复冻融3次,在冰浴中超声碎菌(200W,超声3 s,停4 s,重复99次)。12 000 r/min离心30 min,分别取上清和沉淀进行SDS-PAGE电泳,分析重组蛋白的表达形式。
1.2.7 重组蛋白的纯化 根据优化的表达条件,对阳性克隆进行大量的诱导表达。大量诱导表达后的产物经4℃、12 000 r/min离心10 min后,用细胞裂解液重悬沉淀,反复3次冻融,并冰浴超声裂解细菌,收集上清参照His-镍蛋白纯化套装说明书进行,SDS-PAGE电泳检测纯化的重组蛋白。
2 结果
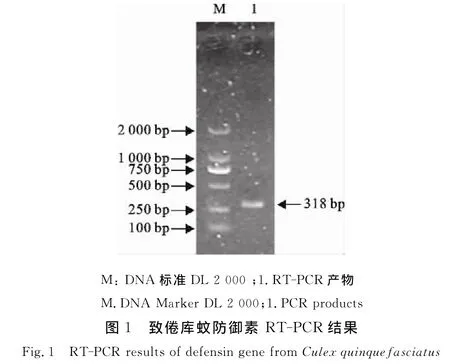
2.1 致倦库蚊防御素基因RT-PCR结果
致倦库蚊总RNA反转录为cDNA后,以Def F和Def R为引物扩增,获得大小约300 bp的特异性扩增条带,与预期大小相符(图1)。测序结果去除两端引物后获得长318 bp片段,含有300 bp完整的开放阅读框,编码99个氨基酸,将此序列登陆Gen-Bank,登录号为JQ799049。
2.2 致倦库蚊防御素成熟肽基因片段PCR扩增结果
以阳性克隆p MD-DEF为模版,PCR扩增产物经10 g/L琼脂糖凝胶电泳检测,结果显示,在250 bp处有一明亮的条带,与预期扩增长度相符(图2)。

2.3 重组质粒pET32a-DEF克隆菌的PCR鉴定
用p ET系列载体的通用T7F/R引物和扩增致倦库蚊防御素成熟肽片段的特异性引物CQF和CQR交叉验证选取的单菌落是否为阳性克隆,并判断目的片段插入的方向是否正确。结果显示,CQF和T7R扩增产物片段约为350 bp,T7F和CQR扩增片段约为750 bp,与预期结果一致(图3),初步证明所挑取菌落为阳性克隆,且目的基因插入载体中的方向正确。
2.4 重组质粒p ET32a-DEF的双酶切鉴定及测序
对空质粒p ET32a(+)和重组质粒p ET32a-DEF分别用Bam HⅠ和Hin dⅢ进行双酶切,结果显示p ET32a-DEF在250 bp出现了一条与PCR产物大小一致的条带,在约6 000 bp处出现了与p ET32a(+)双酶切后大小一致的条带(图4),更进一步判断目的基因成功地与载体连接。测序结果显示重组表达质粒p ET32a-DEF构建成功。


图4 p ET32a-DEF重组质粒的酶切鉴定Fig.4 Identification of recombinant plasmid p ET32a-DEF by enzyme digestion
2.5 重组质粒pET32a-DEF在大肠埃希菌中的表达
SDS-PAGE电泳图可以看出(图5),诱导后的菌体总蛋白比未诱导的菌体总蛋白在相对分子质量约29 ku处可见特异性的蛋白条带,比空载体诱导后表达的蛋白条带大,与预期大小相符。

2.6 IPTG浓度对重组蛋白表达的影响
SDS-PAGE电泳结果显示(图6),IPTG浓度对重组蛋白的表达的影响不明显,用较低浓度的IPTG(终浓度0.2 mmol/L)即可诱导目的蛋白的大量表达。

2.7 诱导时间对重组蛋白表达的影响
SDS-PAGE电泳结果显示(图7),以0.2 mmol/L IPTG浓度进行诱导时,在4 h时表达量较高,继续延长诱导时间重组蛋白表达量基本维持恒定,因而选用4 h为最终诱导时间。

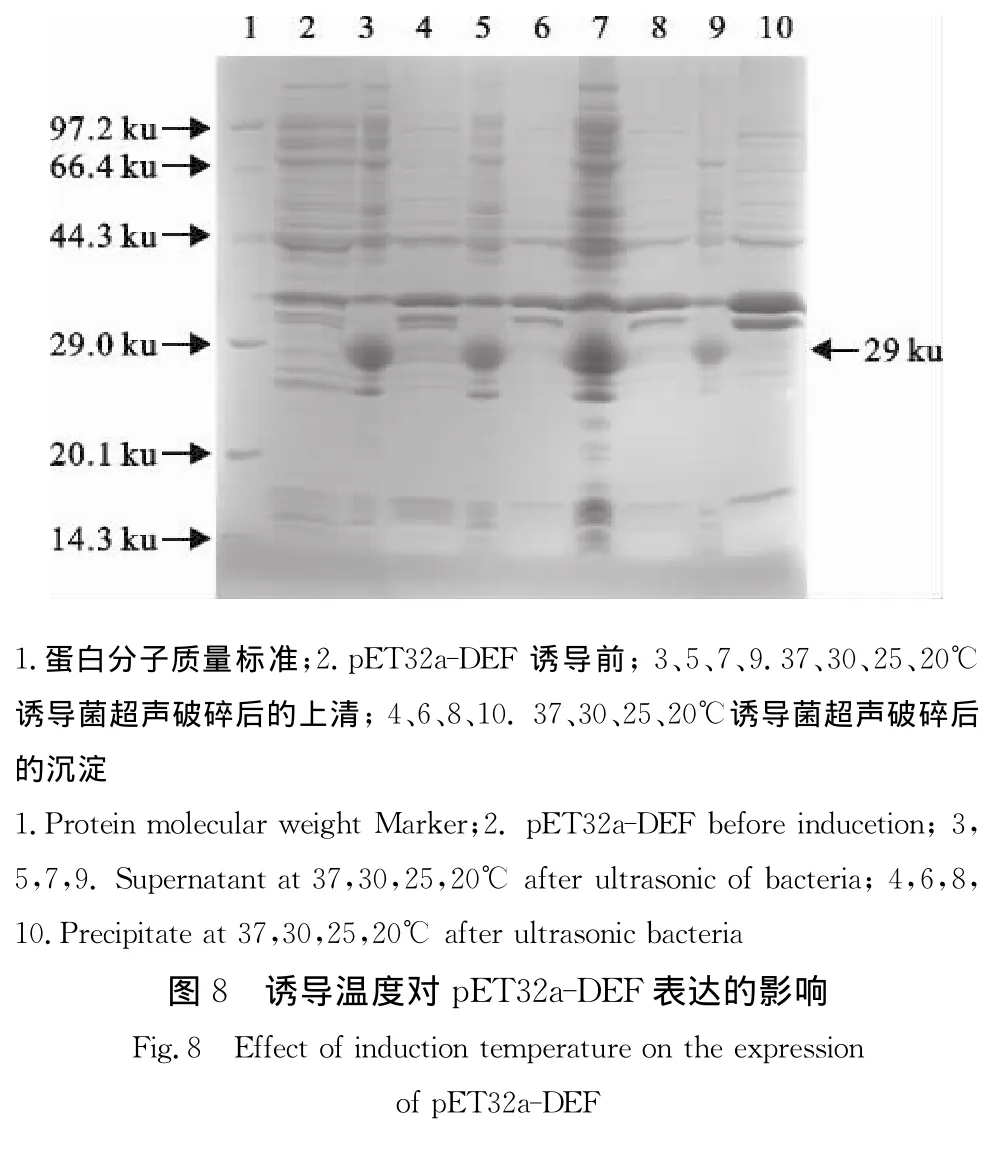
2.8 诱导温度对重组蛋白表达的影响
以0.2 mmol/L IPTG浓度分别在20、25、30、37℃诱导表达4 h后,收集菌体经超声破碎及离心分离上清和沉淀,SDS-PAGE电泳结果显示(图8),重组蛋白在不同温度均存在于上清中。凝胶灰度扫描显示,pET32a-DEF重组蛋白在37℃时上清中的表达量最高,占菌体总蛋白的18.3%。
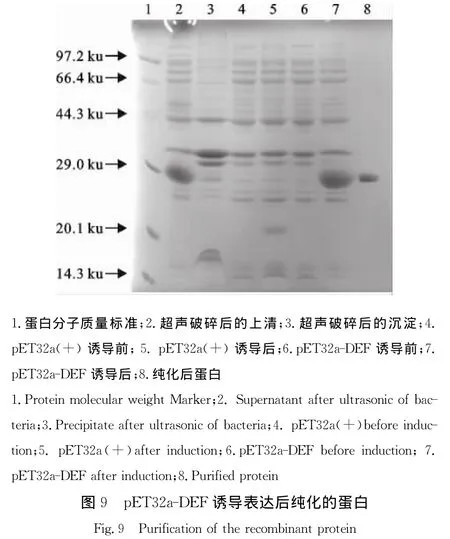
2.9 重组蛋白的纯化
以上述确定的最佳表达条件诱导pET32a-DEF重组蛋白大量表达,超声裂解后将上清按照His– 镍蛋白纯化套装说明进行纯化,SDS-PAGE结果显示纯化蛋白大小约为29 ku,与目的蛋白大小相符,证明蛋白纯化成功(图9)。
3 讨论
昆虫的免疫系统与高等动物相似,也包括阻止异物侵染的体壁、消化道等物理性屏障,体腔内的细胞免疫和多种具有免疫功能的抗菌肽组成的体液免疫[4]。昆虫防御素属于富含半胱氨酸的抗菌肽,其主要抗革兰阳性细菌,偶尔也有抗革兰阴性细菌或真菌的报道[5]。在抗生素抗药性日趋严重的情况下,昆虫防御素极可能成为新的抗菌药物来源。Seufi A M等[6]研究发现夜蛾防御素对革兰阳性和革兰阴性菌均有抑制作用。
致倦库蚊是多种疾病的传播媒介,如乙型脑炎、班氏丝虫病,其幼虫孳生于污染的水体,如粪坑、水坑、水沟等,携带病原体数量多,而自身却并不生病,推测其体内具有强大的防御系统。Kumar B等[7]就曾利用丝虫感染和未感染的致倦库蚊建立差减杂交库,获得了7个与致倦库蚊免疫相关的基因,其中包括防御素基因的部分序列。因而我们认为防御素是致倦库蚊免疫系统中的重要成分,体外获得重组致倦库蚊防御素蛋白,有利于对其功能和作用机理的研究。
利用基因工程技术在大肠埃希菌中高水平表达外源基因,从而获得大量异源蛋白,是现代分子生物学的巨大成就之一[8]。大肠埃希菌作为基因工程表达的首选宿主,具有生产周期短、成本低、产量高等优点[9],能够解决天然资源匮乏、分离纯化困难、化学合成昂贵等问题[10]。
IPTG浓度对外源蛋白在大肠埃希菌中的表达有较大的影响,有时,较低浓度的IPTG即可诱导外源蛋白的大量表达[11]。本试验结果显示IPTG浓度为0.2 mmol/L时即可诱导pET32a-DEF的大量表达,并且增加IPTG的浓度,重组蛋白的表达量没有提高,所以选用IPTG终浓度为0.2 mmol/L作为诱导该蛋白表达的最佳IPTG浓度。诱导时间也是影响外源蛋白表达的一个重要因素,随着诱导时间的延长,外源蛋白表达量将会增加,但是到达一定时间后,外源蛋白表达量将趋于恒定,而杂蛋白的表达量会迅速增加[12]。本研究结果显示当诱导时间到达4 h后,目的蛋白的表达量较高,随时间的推移,重组蛋白的表达量不再增加,因而选择4 h为该蛋白表达的最佳诱导时间。有资料报道,通过低温可以增加重组蛋白的可溶性表达和表达产物的活性,使包涵体的比例下降[13-14]。本研究分别在20、25、30、37℃对重组蛋白进行诱导表达,经超声破碎后分析蛋白的表达形式,结果发现重组蛋白均存在于上清中,以可溶的形式存在,其中在37℃目的蛋白的表达量最高,因而选用37℃为致倦库蚊防御素蛋白表达的最佳温度。
综上所述,本试验通过RT-PCR技术扩增获得致倦库蚊防御素基因全长编码序列,成功构建了致倦库蚊防御素重组表达质粒pET32a-DEF,并对其在大肠埃希菌Rosetta中的表达条件——IPTG浓度、诱导时间、诱导温度进行了优化,获得最佳表达条件为IPTG浓度0.2 mmol/L,诱导时间为4 h,诱导温度为37℃。在此条件下大量诱导目的蛋白的表达,并经His-Ni蛋白纯化柱获得纯化的蛋白,为其功能及作用机理的研究奠定了基础。
[1]Lehre R I,Lichtenstein A K,Ganz T.Defensins-antimicrobial and cytotoxic peptides of mammalian cells[J].Annu Rev Immunol,1993,11:105-128.
[2]周绪霞,陆 平,李卫芬.昆虫防御素及其研究进展[J].免疫学杂志,2004,20(3):121-123.
[3]张亚晶,陈晓光,郑学礼,等.中华按蚊防御素基因cDNA序列和基因组序列的克隆及鉴定[J].中国寄生虫学与寄生虫病杂志,2006,24(1):35-40.
[4]王荫长.昆虫生物化学[M].北京:中国农业出版社,2004:245-270.
[5]高琳琳,李卫东.昆虫抗菌肽及其应用研究进展[J].中国病原生物学杂志,2010,9(5):710-713.
[6]Seufi A M,Hafez E E,Galal F H.Identification,phylogenetic analysis and expression proflile of an anionic insect defensin gene,with antibacterial activity,from bacterial-challenged cotton leafworm,Spodoptera littoralis[J].BMC Mol Biol,2011,11(9):12:47.
[7]Kumar B A,Paily K P.Identification of immune-responsive genes in the mosquito Culex quinquefasciatus infected with the filarial parasite Wuchereria bancrofti[J].Med Vet Entomol,2008,22(4):394-398.
[8]颜文卿,吴德峰,黄印尧,等.犬细小病毒VP2基因原核表达载体的构建与分析[J].福建农林大学学报,2006,35(1):60-62.
[9]陈胜男,马素贞,翟少伦,等.犬细小病毒VP2基因原核表达条件优化与蛋白纯化[J].新疆农业大学学报,2011,34(1):59-61.
[10]张 姝,王 敏,韩梅琳,等.基因重组大肠杆菌表达HrpNEcc蛋白的发酵条件及诱导条件优化[J].中国生物工程杂志,2009,29(10):44-49.
[11]张嘉萱,卢嘉欣,王 蕊,等.人TRAP1基因的克隆、原核表达及表达条件优化[J].生物技术通报,2011(8):161-166.
[12]潘文英,张爱联,张添元,等.用E.coli表达Canstatin-N及其表达条件优化[J].工业微生物,2009,39(1):51-55.
[13]宛传丹,黄宇烽,许晓风,等.人精子蛋白SP22的cDNA克隆、表达及纯化分析[J].中国生物工程杂志,2004,24(12):59-63.
[14]李 琳,潘 英,王正茂,等.丙型肝炎病毒RNA依赖的RNA聚合酶在大肠杆菌中可溶性表达条件的优化[J].药物生物技术,2010,17(3):207-211.
