两品系萼花臂尾轮虫的生活史特征和受精率研究
2012-09-19吕林兰董学兴董斯明蒋琦辰杨家新
吕林兰,董学兴,董斯明,蒋琦辰,杨家新
(1.南京师范大学生命科学学院,江苏南京210046;2.盐城工学院化学与生物工程学院江苏省沿海池塘养殖生态重点实验室,江苏盐城224003)
轮虫是自然水域中分布最为广泛的浮游动物之一,在水域生态系统的物质循环和能量流动中具有重要的作用。由于轮虫栖息地的生态因子 (如水温、光照和食物等)存在差异,不同地理品系之间会产生不同程度的遗传分化,从而在形态或生理生态特征上表现出差异[1-3]。20世纪,研究者发现褶皱臂尾轮虫Brachionus plicatilis是复合体,一些不同地理品系的褶皱臂尾轮虫实际是姐妹种[1]。目前已发现的褶皱臂尾轮虫复合体至少有14个姐妹种,它们不仅在形态、生理生态、遗传等方面存在差异,而且姐妹种之间存在明显的生殖隔离[4-8]。相对于褶皱臂尾轮虫,人们对广泛分布于淡水中的萼花臂尾轮虫Brachionus calyciflorus复合体认识时间不长。李化炳等[9-10]研究发现,萼花臂尾轮虫姐妹种可形成于同一水域,它们的生活史参数具有显著差异。Xiang等[11]研究表明,萼花臂尾轮虫复合体有6个姐妹种,并认为大多数姐妹种的遗传分化与它们的地理距离没有显著的相关性。而Gilbert等[2]研究佛罗里达、佐治亚、德克萨斯和澳大利亚4个地理品系萼花臂尾轮虫的基因相似性和交配行为时发现:遗传差异较大的佛罗里达、德克萨斯和澳大利亚品系间存在生殖隔离现象,这些特性与它们之间的地理分隔相一致。本研究中,作者以萼花臂尾轮虫南京品系和美国佛罗里达品系为研究对象,探讨两品系间和品系内的生活史特征和受精作用差异,以是否形成生殖隔离为指标评价二者是否是姐妹种,旨在为萼花臂尾轮虫的生理生态学及相应领域的研究积累资料。
1 材料与方法
1.1 材料
试验用萼花臂尾轮虫南京品系 (NJ)样品采自江苏省淡水水产研究所养殖池塘(N 32°03',E 118°75');萼花臂尾轮虫佛罗里达品系(FL)休眠卵为美国佐治亚理工学院Terry W Snell教授赠送。
1.2 方法
1.2.1 轮虫的采集与培养 在解剖镜下挑取游泳速度快、个体较大并携非混交卵的雌体,在实验室条件下使用EPA培养液进行单“克隆”培养[12],周期性收集休眠卵保种。以蛋白核小球藻Chlorella pyrenoidosa为饵料进行投喂,小球藻的培养参照杨家新等[13]的方法,投喂前以4 000 r/min离心10 min,再用 EPA培养液将小球藻密度调至 (2~3)×106个/mL。试验在光照培养箱中进行,轮虫培养期间,光照强度为4 000 lx,光照周期L∶D=16∶8,温度为 (25±1)℃。
1.2.2 单个体培养试验 分别挑取单个NJ、FL品系(龄期<4 h)的轮虫幼体放入24孔塑料培养板中,每孔盛新鲜培养液0.8 mL,各设24个平行,小球藻密度为 2×106个/mL,培养条件同“1.2.1”。培养12 h后,每小时观察1次,记录母体和幼体的发育时间(首次携卵)和第一个幼体孵出时间,之后每隔8 h观察1次,记录新生幼体数量,幼体单独培养至携卵,辨别其类型。每隔24 h更换1次培养液,试验至全部母体死亡为止。
1.2.3 生命表构建 分别挑取NJ和FL品系的轮虫幼体 (龄期<4 h)各10个,置于12孔板中,每孔盛新鲜培养液4 mL,各设5个平行,培养条件同“1.2.1”。每隔8 h观察1次,记录各孔轮虫的存活数、死亡数、幼体数,并移去幼体、死亡个体,每隔24 h更换1次培养液,试验至轮虫个体全部死亡为止。
1.2.4 受精率观察 将NJ和FL品系的轮虫进行高密度群体培养,诱导混交雌体,观察到有较多混交雌体出现后,挑取携卵非混交雌体和携雄卵的混交雌体进行单独培养,6 h后,分别挑取非混交卵孵出的幼体和雄卵孵出的雄体。试验共设计4组(NJ♂×NJ♀,FL♂×FL♀,NJ♂×FL♀,FL♂×NJ♀),按雌、雄为1∶3的比例放入96孔板中,每孔加100 μL轮虫培养藻液,24 h后检查并计算受精率,受精率为受精混交雌体 (带休眠卵)占所有混交雌体(带雄卵混交雌体和带休眠卵混交雌体)的百分比。
1.2.5 雌体类型的鉴定
非混交雌体:产非混交卵,个体较大,卵壳较薄而光滑,无色透明且可见胚胎发育过程,在胚胎发育早期可见咀嚼器的几丁质板。
产雄卵的混交雌体:雄卵体积较小,约为非混交卵的1/3,一般带卵数量较多,5个以上。
产休眠卵的混交雌体:经受精后形成,休眠卵个体较大,具有厚的外壳,外表颜色常呈暗褐色,发育成熟的休眠卵两极分化明显[14]。
1.2.6 种群增长参数的计算方法

内禀增长率rm=ln R0/T,
式中:lx(特定年龄存活率)为x年龄组开始时存活个体的百分数;mx(特定年龄繁殖率)为特定年龄出生率,是各年龄组8 h内平均每个雌体的产仔数;rm的精确值是在粗略计算的基础上,根据方程∑lxmxe-rx=1,通过设置程序用试算法在Excel中求得。
应用软件SPSS 16.0对试验数据进行t检验。
2 结果
2.1 两品系萼花臂尾轮虫的幼体发育和胚胎发育历时及繁殖量的比较
试验结果表明:FL和NJ品系幼体的发育时间分别为(12.95±0.22)、(13.47±0.25)h,体外胚胎发育时间分别为(10.00±0.17)、(9.31±0.75)h,第一个幼体孵出时间分别为(22.95±0.28)、(22.78±0.27)h,繁殖量分别为(13.75±0.84)、(16.94±1.10)个,除NJ品系比FL品系的胚胎发育时间显著缩短6.9%(P<0.05)外,两品系间其余指标均无显著差异 (P>0.05)。
2.2 两品系萼花臂尾轮虫的存活率和繁殖率
从图1可见:FL和NJ品系的存活率在48 h后开始下降,NJ品系的存活时间为160 h,FL品系的存活时间为136 h;NJ品系的繁殖率大于FL品系。NJ品系繁殖率>2个/8 h的时间占整个生殖期的50%,其最高繁殖率达到4.1个/8 h;FL品系繁殖率>2个/8 h的时间占整个生殖期的15.38%。
2.3 两品系萼花臂尾轮虫的种群增长参数
从表1可见,NJ品系的净生殖率 (R0)、内禀增长率 (rm)、世代时间 (T)和生命期望 (e0)分别比 FL大 52.88%、12.50%、10.88%和29.08%(P<0.05)。
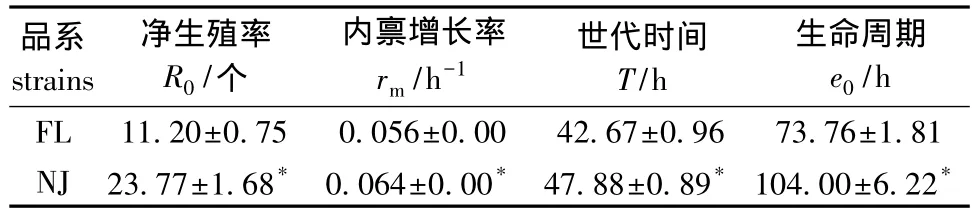
表1 两品系萼花臂尾轮虫的种群增长参数Tab.1 The population growth parameters in two strains of the rotifer B.calyciflorus
2.4 两品系萼花臂尾轮虫的受精率
从图2可见:两品系萼花臂尾轮虫FL和NJ品系内的受精率均为100%,品系间的受精率相对有所下降,其中FL♂×NJ♀的受精率显著低于其他各组 (P<0.05)。

图1 两品系萼花臂尾轮虫的存活率和繁殖率Fig.1 The fecundity and survival rate of amictic female in two strains of the rotifer B.calyciflorus
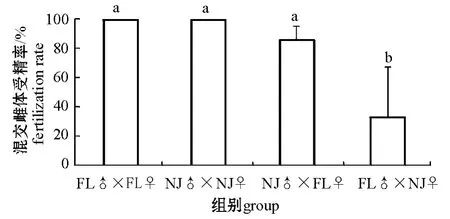
图2 两品系组合萼花臂尾轮虫混交雌体受精率Fig.2 The amictic female fertilization rate between FL and NJ strains
3 讨论
3.1 品系间和品系内萼花臂尾轮虫的生活史特征
已有研究表明,轮虫品系间或姐妹种之间生命表统计学参数差异是由遗传决定的。Ricci等[15]对蛭态类Macrotrachela quadricornifera 5个品系的亮氨酸氨肽酶、精氨酸激酶、葡糖磷酸异构酶、α和β酯酶及苹果酸酶进行分析,发现各品系间有较高的酶基因多态位点,仅有27%的等位基因为各克隆共享,认为各品系生活史的差异不仅是环境诱导的表型可塑性,而且存在基因水平的差异。李化柄等[10]研究了萼花臂尾轮虫3个姐妹种的生命表特征参数,发现姐妹种之间存在类似于镰形臂尾轮虫B.forcatus和尾突臂尾轮虫B.caudatus的种间差异。本研究中得到与上述研究类似的结果,NJ品系和FL品系胚胎发育时间存在显著差异,NJ品系轮虫的净生殖率、内禀增长率、世代时间和生命期望均显著大于FL品系。
3.2 不同品系萼花臂尾轮虫的受精率
轮虫姐妹种的形成机制尚不清楚,地理隔离或栖息地环境因子限制了它们的基因交流可能是形成姐妹种的主要原因[11,16]。而对轮虫姐妹种的判定目前没有统一的标准,形态学、分子生物学方法(线粒体COⅠ、rDNA ITS和16S rDNA序列)是常用的甄别方法[1-2,5,8-9,11,17],而利用轮虫特殊的生活史,观察有性生殖的发生和结果如交配行为、受精率和休眠卵的产生是辨别不同品系是否产生生殖隔离而形成姐妹种的较为简洁而有效的方法[1-2,4,6,9]。已有的研究表明,生殖隔离与遗产差异具有显著的相关性,Fu等[4]研究了7个品系(4个L型,3个S型)褶皱臂尾轮虫的交配试验,发现受精率与交配率与基因距离呈负相关,遗传差异大的L型与S型的受精率为零。Gilbert等[2]研究发现,只有基因水平相近的佛罗里达和佐治亚萼花臂尾轮虫品系能发生交配行为,其他品系之间雄体虽有绕圈行为,但不发生交配,从而认为雄体围绕雌体绕圈的行为与交配行为分别受两个不同的信号控制,并认为地理学与遗传距离具有显著的相关性。而Xiang等[11]以rDNA ITS为标记分析了中国不同地理群体的萼花臂尾轮虫的遗传分化,结果表明,多数姐妹种的遗传分化与温度和pH具有显著的相关性,而与地理学距离没有相关性。另有研究表明,萼花臂尾轮虫和褶皱臂尾轮虫姐妹种可产生于同一水域[9,18]。本试验中以受精率为指标评价地理相隔较远的NJ品系和FL品系萼花臂尾轮虫的生殖隔离状况,发现两者品系间受精率虽较品系内受精率有所下降,但并不存在生殖隔离,由此证明,不同品系萼花臂尾轮虫间是否产生生殖隔离而形成姐妹种与其地理学距离并无相关性。
本研究结果表明,不同品系萼花臂尾轮虫的发育和种群增长参数存在显著差异,但NJ品系和FL品系间不存在完全的生殖隔离,未形成姐妹种。
[1]Gómez A,Snell T W.Sibling species and cryptic speciation in the Brachionus plicatilis species complex(Rotifera)[J].Journal of Evolutionary Biology,1996(9):953-964.
[2]Gilbert J J,Walsh E J.Brachionus calyciflorus is a species complex:mating behavior and genetic differentiation among four geographically isolated strains[J].Hydrobiologia,2005,546:257-265.
[3]冯粒克,席贻龙.三个地理品系萼花臂尾轮虫生活史特征的比较[J].动物学杂志,2004,39(1):12-15.
[4]Fu Y,Hagiwara A,Hirayama K.Crossing between seven strains of the rotifer Brachionus plicatilis[J].Nippon Suisan Gakkaishi,1993,59:2009-2016.
[5]Ciros-Pérez J,Carmona M J,Serra M.On the taxonomy of three sympatric sibling species of the Brachionus plicatilis(Rotifera)complex from Spain,with the description of B.ibericusn sp.[J].Journal of Plankton Research,2001(23):1311-1328.
[6]Gómez A,Serra M.Behavioral reproductive isolation among sympatric strains of Brachionus plicatilis Müller 1786:insights into the status of this taxonomic species[J].Hydrobiologia,1995,313/314:111-119.
[7]Fontaneto D,Giordani I,Melone G,et al.Disentangling the morphological stasis in two rotifer species of the Brachionus plicatilis species complex[J].Hydrobiologia,2007,583:297-307.
[8]Suatoni E,Vicario S,Rice S,et al.An analysis of species boundaries and biogeographic patterns in a cryptic species complex:the rotifer Brachionus plicatilis[J].Molecular Phylogenetics and Evolution,2006,41:86-98.
[9]李化炳,席贻龙,程新峰,等.轮虫同域性物种形成:来自萼花臂尾轮虫克隆间的分子系统发育关系和生殖隔离的证据[J].动物学报,2008,54(2):256-264.
[10]李化炳,席贻龙,程新峰.萼花臂尾轮虫Brachionus calyciflorus种复合体内三个姐妹种生活史特征比较[J].2009,29(2):581-588.
[11]Xiang X L,Xi Y L,Wen X L,et al.Patterns and processes in the genetic differentiation of the Brachionus calyciflorus complex,a passively dispersing freshwater zooplankton[J].Molecular Phylogenetics and Evolution,2011,59:386-398.
[12]United States Environmental Protection Agency(USEP).Methods for measuring the acute toxicity of effluents to freshwater and marine organismsI[C]//U S Environ Protec Agency,Washington D C,1985:216.
[13]杨家新,黄祥飞,刘建康.淡水轮虫繁殖生物学研究进展[J].水产学报,1999,23(3):290-295.
[14]杨家新,黄祥飞.藻类食物对萼花臂尾轮虫繁殖的影响[J].湖泊科学,1998,10(1):42-48.
[15]Ricci C.Comparison of five strains of a parthenogenetic species,Macrotrachela quadricornifera(Rotifera,Bdelloidea)[J].Hydrobiologia,1991,211:147-155.
[16]Gómez A,Carmona M J,Serra M.Ecological factors affecting gene flow in the Brachionus plicatilis complex(Rotifera)[J].Oecologia,1997,111:350-356.
[17]Gómez A,Serra M,Carvalho G R,et al.Speciation in ancient sibling species complexes:evidence from the molecular phylogeny of Brachionus plicatilis(Rotifera)[J].Evolution,2002,56:1431-1444.
[18]Ortells R,Snell T W,Gómez A,et al.Patterns of genetic differentiation in resting egg banks of a rotifer species complex in Spain[J].Arch Hydrobiol,2000,149:529-551.
