疏花水柏枝保护分析与研究
2012-09-11江明喜
江明喜

上:美丽的疏花水柏枝。 摄影/望超
生物多样性保护一直是生态与环境保护的重要内容之一。在《长江三峡水利枢纽初步设计报告(枢纽工程)环境保护篇》和《长江三峡工程生态与环境监测系统实施规划》中,提出了三峡库区需要重点保护的植物有疏花水柏枝、荷叶铁线蕨和川明参。其中,疏花水柏枝分布在库区海拔70-130米的地段,三峡工程于2003年二期蓄水并首次发电后,该区域内生长的疏花水柏枝野生种群已全部被淹没,疏花水柏枝成为唯一自然生境被三峡工程所毁坏的物种。
2007年12月28日,科研人员确认在三峡大坝下游约100公里处的湖北省枝江市董市附近的一处江心洲上发现的植物为疏花水柏枝野生种群,随后在宜都的官洲和宜昌的胭脂坝等江心洲上也发现该物种的存在。在三峡大坝下游的3个江心洲上发现疏花水柏枝新的分布区,对于该物种的就地保护具有重要意义。
三峡库区疏花水柏枝种群的基础研究
对疏花水柏枝的研究和保护主要源于三峡工程的建设,生态环境保护问题受到空前的重视,疏花水柏枝的保护同样受到相当的关注,相关部门制定了保护规划,对疏花水柏枝等一批物种进行了迁地保护和专题研究。疏花水柏枝的研究可归纳为以下几个方面:
疏花水柏枝的分类地位
疏花水柏枝的第一份标本是A.Davidi采自湖北巴东一带的长江谷地,1888年,A.R.Franchet根据这份标本,将其定为水柏枝的一个变种;1984年,我国植物分类学家将其提升为种,命名为疏花水柏枝(张鹏云等,1984),是我国柽柳科水柏枝属分布最东的一个种。疏花水柏枝将因三峡工程建设而导致生境丧失。因此,在《中国红色物种名录》(第一卷)中,疏花水柏枝被列为濒危种。
疏花水柏枝高0.6-1.0米,少数高达1.5米。叶片鳞片状,紧贴枝干,上部微有外翘。花白色至微粉红色,花梗长2毫米,花萼深5裂,花瓣5枚,雄蕊10,长短相间排列,花丝下部合生,雌蕊3心皮组成,蒴果狭圆锥形。花序为穗状花序,每年夏天6月开花,可持续到10月。果为蒴果,种子具毛。疏花水柏枝根系发达,长0.3-1.5米不等,根尖出露地表,也可重新长出枝条,形成新的植株,有的老枝条被沙压住后能形成新的植株。
每年夏天6-7月长江丰水期,疏花水柏枝一般被全部淹没,这段时间也是它落叶(或枯叶)期,水退后,花序仍不断开花,完成结果及种子散布。
围绕疏花水柏枝开展的基础科研工作
我国对该物种开展了一系列的包括种子结构、群落生态、繁殖技术、种群动态、遗传多样性和生理生态等方面的研究工作。陈伟烈等(1994)通过调查,确定了该物种的地理分布范围,并首先预见了三峡工程修建后会全部淹没该物种的全部生境,使之成为一种濒危物种。吴金清等(1998)对疏花水柏枝野外种群与群落组成与结构、种群分布生态环境进行了研究。疏花水柏枝的扦插繁殖也得到了研究(徐慧珠等,1998;熊高明1996;韩敏等,2008)。李作洲等(2003)采用超薄平板微型聚丙烯酚胺等电聚焦电泳方法对疏花水柏枝的13个自然居群和1个人工迁地保护居群的等位酶遗传变异进行了初步研究,检测了5个酶系统,得到13个等位酶位点,认为疏花水柏枝具有较高水平的遗传多样性,在分析讨论疏花水柏枝遗传多样性与其繁育系统、生境及其起源进化的关系的基础上,探讨了疏花水柏枝濒危的主要原因,推断其应为第四纪冰川影响后的古孓遗种。刘义飞等(2006)等通过对三峡库区的疏花水柏枝的遗传多样性进行分析,发现该种基因流为单向流动,即从上游向下游的扩散格局。王勇等(2003)对疏花水柏枝的地理分布、自然生境和群落结构进行了系统调查,发现疏花水柏枝分布于三峡库区12个县级单位的长江干流消涨带,共31个居群9万余株,比原己知的分布区增加了7个县级区域18个居群8万余株。并认为该物种具有喜温湿、耐水淹和生长反季的生长习性,具有种子多、易扦插的繁殖优势和自然居群中物种少、水淹前后物种组成有差异的群落结构特点,并提出了一些相应的保护意见。陶勇等(2004)以疏花水柏枝茎的11个解剖结构参数作为表型性状,对覆盖整个分布区域的居群进行部分抽样取样,研究了该物种表型性状的多样性及其格局、表型性状的分化及其与环境的关系,认为各性状在居群间以及不同居群总体性状之间都存在显著或极显著的差异。对其种子形态解剖研究表明成熟种子具直立型胚,无胚乳,种皮细胞一层,其外围有较厚的脂质层。胚轴和子叶的细胞均纵列排列,具较大列间隙,有时在胚轴上可见位置不定的坑。胚根区有栅栏状细胞,珠孔区以上有薄壁细胞解体后形成的断裂层。中央维管网贯穿胚体,其盲端区常出现不能与之连通的局部输导组织。种子的结构特征为水流扩散提供了便利,但也限制了扩散效力(陶勇等,2008)。陈芳清等(2005a)对疏花水柏枝幼苗密度与生长的关系进行了研究,并对迁地保护的种群重建提出了自己的看法(陈芳清等,2005b)。陈芳清等(2008)对疏花水柏枝的光合作用、蒸腾作用、水势等生理特性以及丙二醛(MDA)、过氧化物酶(POD)和多酚氧化酶(PPO)活性等进行分析测定。
已开展的迁地保护工作
对疏花水柏枝的保护工作在上个世纪90年代初就已开始,主要是开展扦插繁殖、迁地保护、离体培养和种子超低温保存等方面的工作。到目前为止,中国科学院武汉植物园和中国科学院植物研究所的科研人员在野外和植物园建立了7个迁地保护点,分布在湖北省兴山县南阳镇凉水井香溪河河滩地、重庆巫溪县花台乡花台村鲤鱼山大宁河河滩地、重庆丰都县世坪森林公园、宜昌市长江大桥南岸江滩地、秭归泗溪、宜昌市三峡植物园(金银岗)、中国科学院武汉植物园(武昌磨山),这些地点共保存植株7000株。
在实验室开展的设施保存实验中,-18℃(冷冻)的温度条件下存放低温保存的种子发芽率均超过80%,表明保存的条件基本达到了疏花水柏枝种子低温保存的最适条件。离体保存的继代培养则不理想,表现为继代培养1-2代均生长良好,但3代以后存活率则急剧下降,无法满足长期保存的要求。
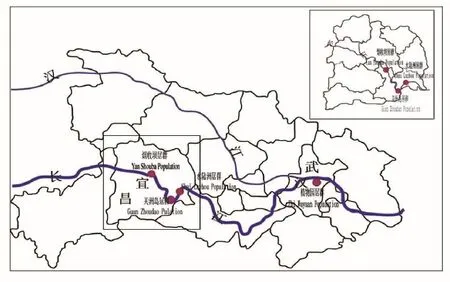
图1 疏花水柏枝实验居群分布图
新发现残存疏花水柏枝种群的现状
新发现的野生疏花水柏枝种群主要分布在长江宜昌河段胭脂坝岛和宜都市关洲岛(30°39.270′N,111°18.958′E),而枝江董市小岛上的数量较少(见图1)。该地区属亚热带季风性湿润气候。四季分明,春秋较长。年平均降水量为992.1~1404.1毫米之间。雨水丰沛,多在夏季,较长的降水过程多发生在6~7月份,雨热同季,全年积温较高,无霜期较长,约300天。年平均气温为13.1~18℃,极端最高气温41.4℃,最低气温-15.6℃。
每年5-9月由于上游大坝排洪,江心洲大部分面积被淹没。两地距离约70公里,生境和植被类型比较相似。生境类型主要为砾石基质和河沙基质,相较而言,胭脂坝岛内砾石基质分布更多。由于长期受到长江汛期水淹干扰,植被以灌丛为主,且群落的物种组成也比较简单。主要的物种有:秋华柳、疏花水柏枝、狗牙根、苔草、灰绿藜、萹蓄、繁缕等。而分别对两岛进行群落学调查显示,关洲岛疏花水柏枝种群比在胭脂坝岛分布更广,数量也更多。
根据调查估算,关洲岛上的个体数量约20000株。该岛交通不便,保存状况良好。胭脂坝位于宜昌市区和点军之间的长江江心洲上,正在建设中的宜万铁路的过江高架桥横跨而过,这里曾经是修桥工人的居住地,在岛的前端宜昌市政府在修建江心洲沙滩的护沙工程,附近的一些工程建设也到该岛上取细纱石料等,所以,该岛的人类活动十分强烈。该岛上的疏花水柏枝的生境比较单一,全部是鸡蛋大小的砾石为基质,由于取细沙石料等原因,疏花水柏枝的分布明显聚集成片状。与关洲岛相似的是,该岛上4年生左右的苗木很多。根据调查,该岛上的种群数量约20000株。枝江董市的江心洲已承包给一民营企业家开发沙滩旅游,该岛上疏花水柏枝种群数量约40株。
疏花水柏枝传粉生物学研究
疏花水柏枝残存居群和迁地保护居群传粉生殖进程的变化
疏花水柏枝为多年生灌木,野生残存居群植株在5至9月处于水淹休眠状态,9月末江水下落后,江心岛上的居群露出水面后,迅速恢复生长,经过约1-2星期的营养生长后,部分植株进入花期。10月中旬至11月下旬,绝大部分植株进入花期。12月初气温下降,传粉者活动减少,花期逐渐结束。武汉植物园迁地保护居群花期为6月至12月,但夏季7月至8月有性生殖失败(图2);疏花水柏枝具无限花序,为同步大量开花模式。居群内大部分植株几乎同期进入花期,并且在植株水平上,各枝条上的花序同样表现出高度的同步性。花自基部向顶部开放,平均每天开2-3朵,单个花序的花经历30-60天全部开完。雌雄同熟,单花寿命1-2天。授粉后的小花子房迅速膨大,约20天左右成熟,即开裂散播种子。种子由风力或水力传播,遇到适宜的环境即可在24小时内萌发。所以,由花序基部向顶端,种子散布、果实成熟和小花开放同时进行。
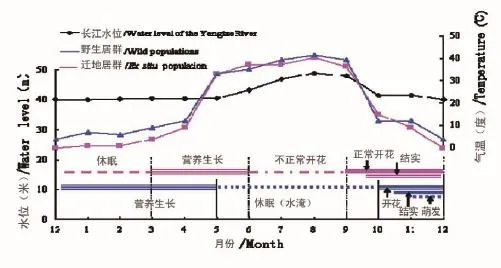
图2 不同生境条件下疏花水柏枝的生长周期
疏花水柏枝的繁育系统及不同保护环境下传粉昆虫的变化
疏花水柏枝为以异交为主,部分自交亲和的虫媒繁育系统。人工套袋授粉实验的结果发现:去雄套袋不结实,表明疏花水柏枝不存在无融合生殖;不去雄套袋也不结实,表明疏花水柏枝不存在自动自交现象;去雄套120目网袋(网孔大小可以使花粉通过,但访花昆虫不能通过)不结实,表明不存在风媒传粉。疏花水柏枝在自然情况下的结实率为0.86±0.07(n=120)。除去雄套袋、不去雄套袋和去雄套网袋3种处理外(因为这三种处理均不结实,无法进行单因素方差分析),经过one-way ANOVA方法分析,各处理组结实率差异极显著(p<0.01),自然结实显著高于自交和异交处理,并且异交处理显著高于自交处理。
在疏花水柏枝野生残存居群内观察发现,其访花昆虫主要是膜翅目、双翅目和鳞翅目类昆虫,包括12个科20种。主要传粉者是双翅目的长尾管食蚜蝇、黑色锥口蝇、大头金蝇、家蝇和膜翅目的中华蜜蜂、意大利蜜蜂。在武汉植物园迁地保护居群中的传粉昆虫主要也是上述3目10余种昆虫,主要变化是在迁地居群中发现在残存居群中不存在的蚂蚁传粉,导致更多的近交繁殖。
对传粉者单花滞留时间、单花间飞行时间、单花传粉时间(即单花内滞留与飞行到下一朵花的时间之和)和花序滞留时间进行one-way ANOVA方法分析,结果表明:不同传粉者单花滞留时间差异显著,意大利蜜蜂单花滞留时间(1.44±0.28s)显著低于大头金蝇(4.14±0.63s)、长尾管蚜蝇(3.99±0.62s)和中华蜜蜂(2.85±0.23s);单花间飞行时间和单花传粉时间差异不显著(图3)。相比之下,意大利蜜蜂的单花间飞行时间(1.71±0.10)要大于其它传粉者,但没有达到显著性水平;黑色锥口蝇的单花传粉时间(4.96±1.66)略高与其它传粉者,同样也没有达到显著性水平。花序滞留时间差异不显著,但两两比较得出:大头金蝇花序访花时间(27.43±4.54s)显著高于意大利蜜蜂(7.02±3.95s),同其它传粉者差异不显著(图3)。
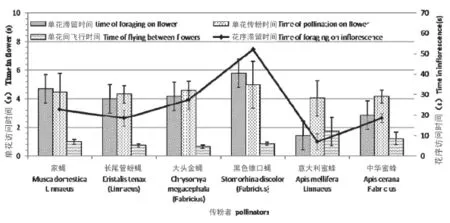
图3 疏花水柏枝主要传粉者访花行为

2008年12月22日,湖北宜昌市城区长江南岸胭脂坝江滩上的三峡特有植物疏花水柏枝长势良好(疏花水柏枝又称为“植物大熊猫”)。摄影/刘君凤/CFP
疏花水柏枝遗传多样性研究
研究采用10条ISSR引物对343个疏花水柏枝个体进行了扩增得到147条带谱,其中141条具有多态性。没有出现单个居群特有带谱,但是当以三峡大坝为界将12个居群分为两个组时,在库区被淹没的居群中存在3条特有带谱,而在残留居群中存在1条特有带谱。研究结果表明,疏花水柏枝在物种水平(PPB=95.9%)和居群水平(PPB=58.5%~80.3%)都表现出高的ISSR遗传多样性。在研究的12个自然居群中,宜昌居群和宜都居群具有较高的遗传多样性水平,而枝江居群的遗传多样性最低。疏花水柏枝高的遗传多样性水平与基于同工酶和AFLP的研究报道结果是相符的。繁育系统、地理分布范围以及扩散模式是导致疏花水柏枝具有高水平遗传多样性的主要原因。
以所有扩增得到的147条带谱表示疏花水柏枝携带的遗传信息,3个残留居群涵盖了该物种绝大部分的遗传信息(97.96%),尤其是宜昌居群97.96%)和宜都居群(91.84%),而枝江居群涵盖的物种遗传信息相对较少(71.43%)。枝江居群表现出与其它11个居群间具有很大的遗传差异,而宜昌居群和宜都居群与库区被淹没的9个自然居群具有较近的遗传关系。我们推测疏花水柏枝3个残留居群在三峡库区居群被淹没之前就已经存在,这三个居群可能来源于该物种早期沿长江流域的扩散,而且枝江居群可能由于河道的变化等原因,导致该居群与其他居群的基金交流受限,使该居群与其他居群发生了分化。但是,由于ISSR分子标记的局限性,对于残留居群的起源和历史还需要通过保守性较高的叶绿体SSR或基因组序列等分子方法进一步研究证明。
疏花水柏枝的保护对策
物种保护的途径主要有就地保护和迁地保护,残留疏花水柏枝居群的存在使得该物种的就地保护成为可能。残留居群中的宜昌居群和宜都居群在研究的12个居群中具有最高的遗传多样性并产生了这两个居群特有的等位基因,而枝江居群的遗传多样性虽然较低,但是在野外观察中该居群内幼苗更新有限,因此,迫切需要对3个残留居群进行保护,尤其是在库区自然居群全部消失的情况下。在野外观察中宜昌居群和宜都居群具有很强的幼苗更新能力,因此建设自然保护小区对这两个居群可以进行就地保护。而枝江居群中表现出的后代适合度下降,使得对该居群进行就地保护的同时有必要进行迁地保护。在对3个残留居群进行保护的同时,对已有的迁地居群也要进行合理的管理以尽可能减少该物种遗传资源的流失。
三峡工程建成后改变了下游原有水文节律,清水下泻将对残留疏花水柏枝的生境带来影响,这种影响对该物种的长期生存意味着什么?一切需要长期的定位监测才能回答。
