鸡痘病毒分离株及疫苗株整合网状内皮组织增生病病毒的基因分析
2012-09-11车国喜孙洪磊刘立涛刘思当
车国喜,孙洪磊,于 宏,李 超,刘立涛,刘思当*
(1.山东农业大学动物科技学院,山东泰安271018;2.中国农业大学,北京100193;3.泰安市畜牧兽医局,山东泰安271018;4.夏津县畜牧兽医局,山东夏津253200)
禽网状内皮组织增生病病毒(Reticuloendotheliosis virus,REV)基因片段或全基因整合进鸡痘病毒(Fow lpox virus,FPV)已成为关注的焦点。国外陆续报道了FPV整合有REV前病毒基因序列,即野毒株中含有REV病毒全基因序列,疫苗中整合有REV的长末端重复序列(Long terminal repeat,LTR)片段或全基因序列[1-6]。在我国,自2004年丁家波、崔治中等首次报道FPV基因组中整合有REV基因[7]以来,近几年陆续有报道称分离到的FPV野毒株中含有REV前病毒全基因序列,疫苗株整合有REV的LTR片段或者是全基因序列[8-9]。FPV基因组中有完整REV前病毒基因组整合对养鸡业是一种的潜在危胁[5]。至今,FPV野毒株的REV整合区全基因分析以及有关FPV野毒株和疫苗株整合序列差异比较的报道尚属少见。因此,本研究通过基因分析,了解山东地区FPV野毒流行株整合REV序列的情况,同时比较FPV分离株和弱毒疫苗株自然整合REV前病毒序列的差异,评估国内当前使用疫苗的缺陷及潜在风险,进而为有效预防鸡痘及禽网状内皮组织增生病奠提供实验依据。
1 材料和方法
1.1 病毒株及主要试剂 14个FPV分离株为山东农业大学动科院临床病理学实验室于2008年~2011年间自山东各地送检的鸡痘典型病例中分离鉴定及保存,代号依次为1~14;5株疫苗代号依次为:V1、V2、V3、V4和V5,其中V5为进口疫苗株。LA Taq DNA聚合酶、r Taq、DL15000DNA Marker、胶回收试剂盒等购自TaKaRa公司。
1.2 实验动物及病毒扩增 9日龄~11日龄SPF鸡胚购自山东省农科院家禽所;痘病毒增殖按常规接种鸡胚的方法进行。
1.3 引物设计与合成 根据普遍整合位点,参考文献[1]设计合成两对扩增FPV整合区全长序列引物(表 1)。
1.4 PCR反应条件及产物纯化 P1/P2引物:93℃1min、55℃ 2min、68℃ 6.5m in,30个循环。P3/P4引物:93℃1m in、55℃2m in、68℃6m in,30个循环。201F/203R引物:用r Taq酶扩增:94℃1m in、56℃ 1min,72℃ 6.5min,30个循环。用LA Taq酶扩增:94℃50s、56℃30s、72℃9m in,30个循环。
PCR产物经琼脂糖凝胶电泳检测,并采用胶回收试剂盒回收、克隆于pMD18-T载体中由上海生工生物工程技术服务有限公司测序。

表1 引物序列Table 1Sequence of primers
1.5 测序及基因分析 对5个FPV疫苗株和随机挑选的4个分离株进行测序,测序结果采用DNA Star软件进行序列拼接、比对、同源性分析并生成基因进化树。
2 结果
2.1 FPV疫苗株及分离株的REV整合区PCR扩增结果 以疫苗直接提取的DNA为模板,采用特异性引物对201F/203R进行扩增。分离株以感染的鸡胚鸡胚绒毛尿囊膜(CAM)上的痘斑研磨液提取的DNA为模板,分别采用特异性引物对P1/P2和P3/P4进行扩增。扩增产物经琼脂糖凝胶电泳检测,结果表明4株国产疫苗扩增片段大小相近(图2A),约1400bp,而进口疫苗V5片段较大,约1700bp;14个分离株扩增片段大小基本相同,P1/P2引物对和P3/P4引物对均得到了预期大小分别为6300bp(图 2B)和 5900bp(图 2C)的片段。
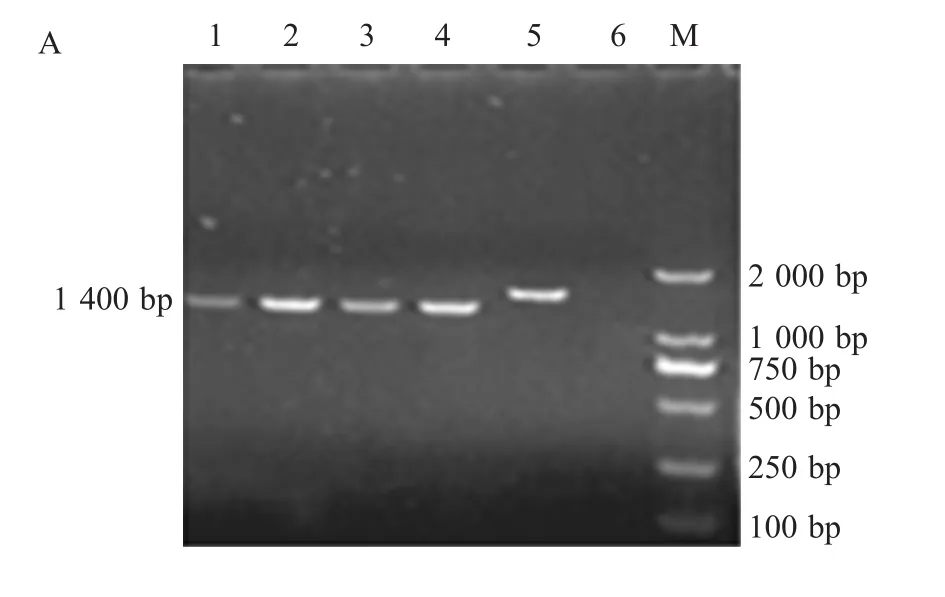
A:PCR amplification of FPV vaccine strainswith 201F/203R primers;1-5:PCR products of V1,V2,V3,V4and V5;6:Negative control;M:DNA Marker
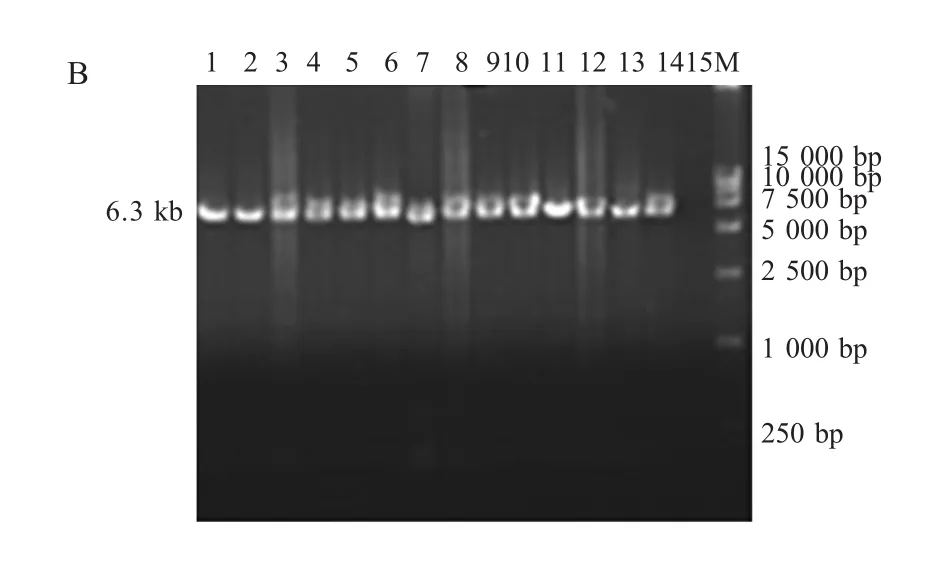
B:PCR amplification of FPV isolateswith P1/P2primers;1-14:PCR products of FPV isolates 1-14;15:Negative control;M:DNA Marker
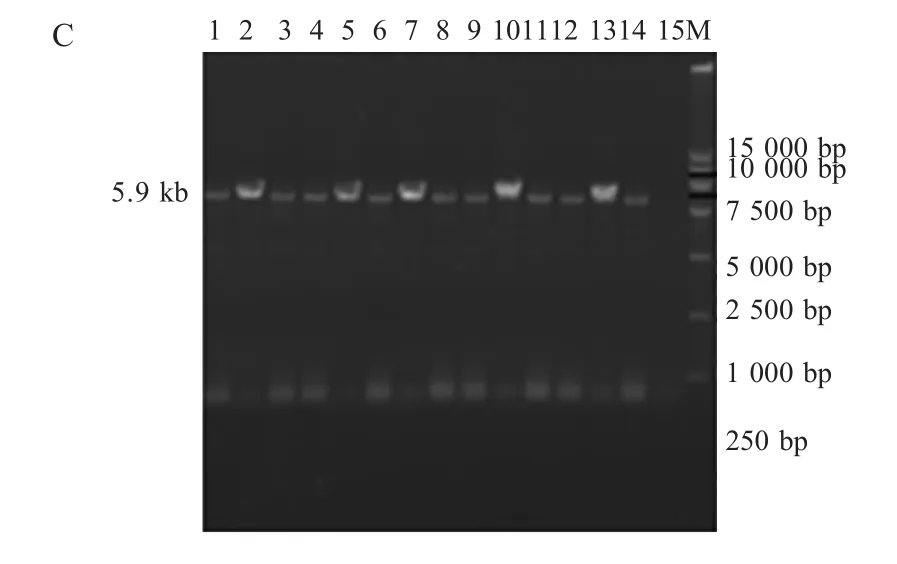
图1 FPV疫苗株及分离株REV整合区PCR扩增结果Fig.1Amplification of REV integrated regions of FPV vaccine strains and isolates by PCR
2.2 基因分析
2.2.1FPV疫苗株的整合区基因分析测序结果表明4株国产疫苗扩增序列长度分别为1378bp、1381bp、1382bp和1381bp,将测序结果录入到GenBank中进行Blast显示,4个序列均由3部分组成,aa 1~aa 822均为FPV的ORF201基因编码,长822bp;第二部分均为REV的5'LTR,位于823bp~1015bp,长193bp;第3部分为FPV的ORF203,长分别 363bp、366bp、367bp和 366bp;这 4个整合LTR序列同源性高达100%,与FPV株AF006064(澳大利亚疫苗株)、AF198100(强毒株)、AY255633(分离株)和HP-438(高代次分离株)的同源性高达99.9%~100%。表明REV-LTR在FPV基因组中的存在具有很悠久的渊源。
但整合的LTR序列与REV的美国标准株SNV和中国标准株HA9901同源性分别为70.4%和76.3%(表 2),与国外(APC-566)、台湾(Chicken 3337/05和Goose 3410/06)、中国大陆(HLJ071、ZD0708和HLJR0901)的REV分离株同源性均为99.0%。与国外分离的整合REV全序列的FPV野毒株(AF246698)的LTR序列同源性平均为100%(表2),表明这些LTR整合序列与REV株的LTR序列同源性不尽相同,与REV标准株同源性较低,与近期分离的REV株LTR序列的同源性较高,而与已知的整合株LTR同源性则高达100%。基因遗传进化树分析也显示了这种关系(图2)。
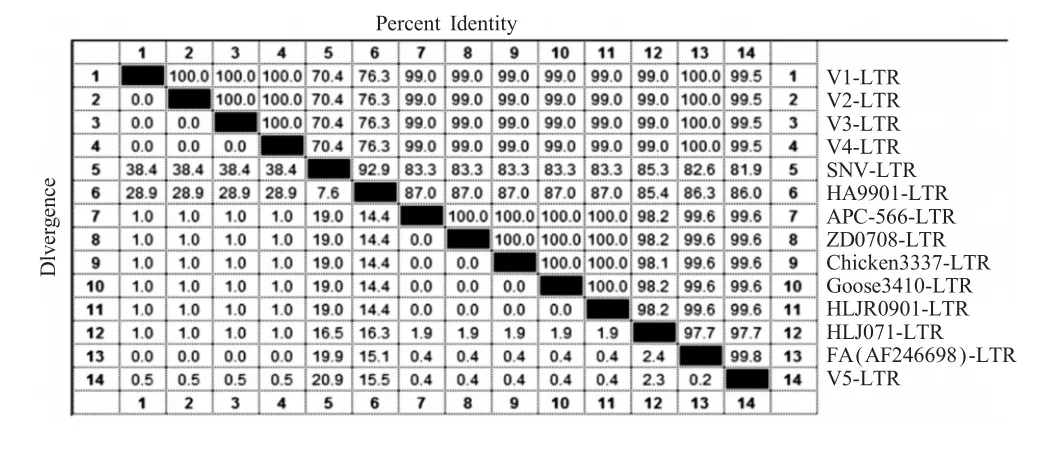
表2 整合LTR序列的同源性图谱Table 2Homologymap of integrated LTR
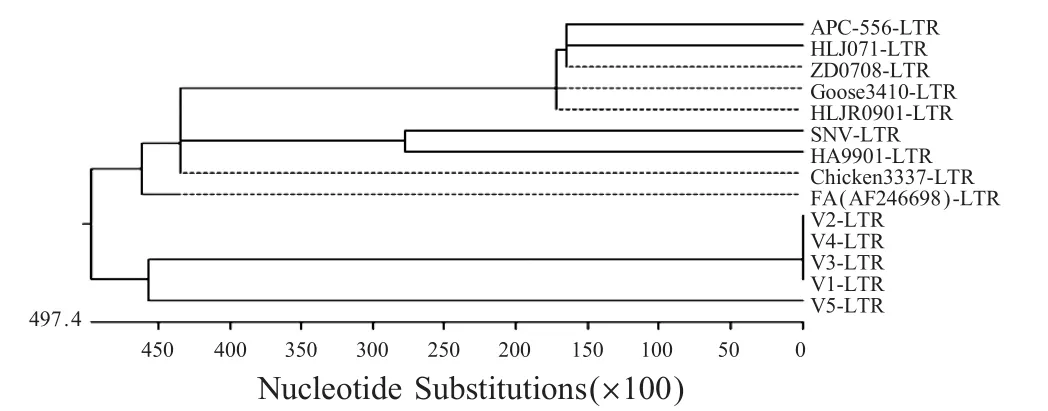
图2 FPV疫苗株整合LTR基因进化树Fig.2 Genetic evolution tree of integrated LTR for FPV vaccine strains
然而,进口疫苗V5的片段长度为1638bp,其中整合的LTR长度为486bp,该LTR序列与REV标准株和已知REV株的同源性与4株国产疫苗相似,但与国外分离株的整合全长LTR序列的FPV AY255633的LTR序列同源性高达100%,与4株国产疫苗相比,进口疫苗V5整合的LTR片段相对较完整。
2.2.2FPV分离株的整合区基因分析随机选取的4个分离株5、10、12和14进行测序,拼接之后长度分别为10306bp、10306bp、10168bp和10304bp,这些序列均由3部分组成:第1部分为FPV ORF201的1bp~904bp,各株间相同;中间部分为整合的REV序列,5、10、12和14株该段长度分别为7938bp、7938bp、7799bp和 7936bp;第3部分为FPV的ORF203,长度1464bp。样品间整合区REV序列同源性为99.6%~100%(表3),除分离株12(YY2010)在REV的gag基因有两段共138bp的缺失以外,其间差异甚微。
这些整合序列与美国分离的整合株整合区REV序列同源性最高,均在99.5%以上;与美国REV标准株SNV同源性平均为96.1%(95.9%~96.2%之间),与国内REV标准株HA9901同源性平均为97.7%(97.4%~97.8%之间),而与国内外分离的REV株的同源性差异不大,均在99%以上(表3)。以上结果表明山东地区的FPV分离株的整合区REV序列高度同源,与国外FPV整合株的整合区REV序列同源性最高,与国内外REV分离株的同源性较高并且差异不大,而与标准株的同源性较低。基因遗传进化树分析也显示了这种关系(图3)。
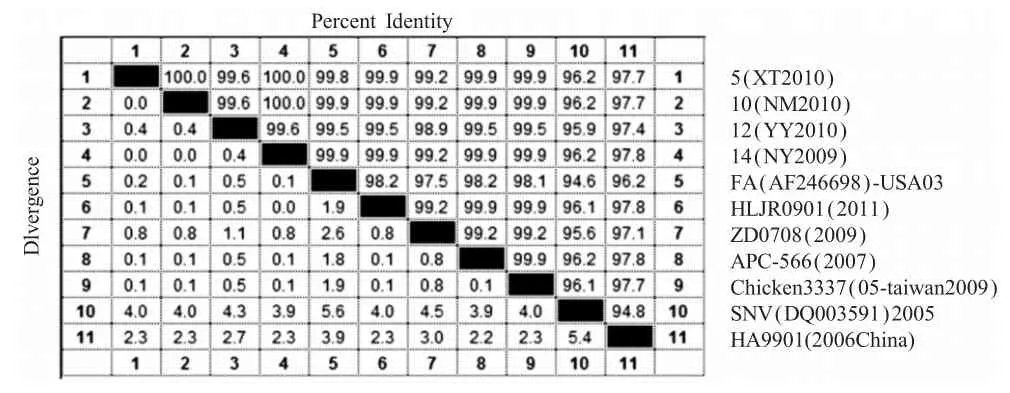
表3 FPV野毒株整合区基因的同源性图谱Table 3Homology of integrated sequences from FPV field strains

图3 FPV分离株整合区基因进化树Fig.3Genetic evolution tree of integrated sequences from FPV isolates
整合区LTR序列比较结果显示:4个分离株整合的LTR序列基本一致,5'LTR长度为517bp,3'LTR长度为222bp,与国内外已知的REV株相比,这些整合株的3'LTR变异较大,其长度明显缩短。
3 讨 论
REV序列在FPV疫苗株和野毒株中的整合已是普遍现象,但对于整合序列的报道并不多见。本研究根据普遍存在的整合位点,分别从FPV和REV基因设计上下游引物,在得到整合区全序列的同时,还避免了因REV游离病毒粒子存在导致的非特异性扩增。
本研究结果显示2008年~2011年间分离的山东地区14个FPV分离株整合REV前病毒的位置一致,均位于FPV-201ORF和203ORF之间,而且极其一致地整合了几乎完整的REV前病毒基因序列。整合的REV序列与国外分离的整合株同源性最高,与REV株同源性较高,但与REV标准株同源性较低。其中整合区LTR序列变异最大,尤其是3'LTR明显缩短,这表明整合的REV序列经过了修饰,使之与FPV序列之间的结合更适于两者的生存和复制,整合过程中的这种修饰与REV-LTR,尤其是3'LTR的明显缺失有关。国外也有类似报道[3-4,9]。分离株5、10和14的整合区序列高度同源,无论是整合的LTR基因还是中间的gag、pol和env基因均差异甚微,这有可能如现在报道的两株MDV所体现出来的现象[10],即这3株FPV野毒株很有可能是来自同一个原始重组病毒,由该原始重组病毒在鸡群中形成的流行病毒株。而分离株12由于gag基因出现较区域缺失可能与其他3株来自不同的原始重组病毒。
然而,国内外不同厂家的5株疫苗,均仅仅整合了REV前病毒的5'LTR序列,这与国外报道的所有FPV疫苗中均存在REV-LTR序列一致[4]。这些LTR序列在长度上分为两种,国产疫苗整合的LTR长度相对于进口疫苗较小。对于疫苗整合片段大小不同,在疫苗效力、安全性等方面的差异有待进一步研究。
整合几乎全长REV前病毒序列的FPV已成为流行鸡痘的优势病毒株,关于整合野毒株的致病特性,国外有报道称整合的REV序列具有感染性,并且可以复制,同时引起感染鸡出现持续3周以上的病毒血症,并表现明显的免疫抑制作用[1-2,6]。本实验室前期的研究中显示FPV整合野毒株除表现痘病病变外,还表现某些REV的致病特性,造成感染鸡持续性的免疫抑制,使REV抗体转阳,并出现REV病毒血症[11-12],进一步证实REV可以通过基因组整合于FPV而进行传播的推断。
但目前鸡痘的高发病率和免疫失败现象与REV前病毒序列整合相关的更多内在联系,整合部分或全部REV-LTR序列的FPV疫苗的安全性,以及这些疫苗对整合几乎全长REV前病毒序列的FPV野毒的免疫保护作用,目前未见深入的研究报道,应进一步研究探讨。
[1]Heritig C,Coupar B E,Gould A R,et al.Field and vaccine strains of fow lpox carry integrated sequences from the avian retrovirus,reticuloendotheliosis virus[J].Virology,1997,235:367-376.
[2]Singh P,Kim T J,Tripathy D N.Re-emerging fow lpox:evaluation of isolates from vaccinated flocks[J].Avian Pathol,2000,9(5):449-455.
[3]Tadese T,Reed W M.Detection of specific reticuloendotheliosis virus sequence and protein from REV-integrated fow lpox virus strains[J].JVirol Methods,2003,110:99-104.
[4]Fadly A,Garcia M C.Detection of reticuloendotheliosis virus in live virus vaccines of poultry[J].Devel Biol,2006,126:301-305.
[5]Wang Jian-ning,Meers J,Spradbrow P B,et al.Evaluation of immune effects of fow lpox vaccine strains and field isolates[J].Vet M icrobiol,2006,25;116(1/3):106-119.
[6]Singh P,Schnitzlein W M,Tripathy D N.Construction and characterization of a fow lpox virus field isolate whose genome lacks reticuloendotheliosis provirus nucleotide sequences[J].Avian Dis,2005,49(3):401-408.
[7]丁家波,崔治中,于立娟,等.含有禽网状内皮组织增生病病毒基因组片断天然重组禽痘病毒的研究[J].微生物学报,2004,44(5):588-592.
[8]于立娟,崔治中.禽痘疫苗病毒中网状内皮组织增生病病毒5'LTR整合位点序列分析[J].微生物学报,2006,46(4):660-662.
[9]陈静.禽痘病毒基因组中整合有禽网状内皮组织增生病病毒序列的研究[D].扬州:扬州大学,2006.
[10]崔治中.禽反转录病毒与DNA病毒间的基因重组及其流行病学意义[J].病毒学报,2006,(22):151-153.
[11]王刚,孙洪磊,于颖,等.鸡痘病毒整合禽网状内皮组织增生病病毒基因的分子生物学鉴定及致病机理研究[J].中国农业科学,2009,42(11):4077-4084.
[12]孙洪磊,车国喜,秦梅,等.整合禽网状内皮组织增生病病毒基因的鸡痘病毒野毒株致病性研究[J].中国兽医学报,2011,31(11):1568-1572.
