濒危植物大果木莲种群格局及濒危原因分析
2012-09-06陈少瑜付玉嫔吴涛司马永康郝佳波韩燕
陈少瑜,付玉嫔,吴涛,司马永康,郝佳波,韩燕
(1.云南省林业科学院国家林业局云南珍稀濒特森林植物保护和繁育重点实验室云南省森林植物培育与开发利用重点实验室,云南 昆明 650204;2.西南林业大学资源学院,云南 昆明 650224)
濒危植物大果木莲种群格局及濒危原因分析
陈少瑜1,付玉嫔1,吴涛1,司马永康1,郝佳波1,韩燕2
(1.云南省林业科学院国家林业局云南珍稀濒特森林植物保护和繁育重点实验室云南省森林植物培育与开发利用重点实验室,云南 昆明 650204;2.西南林业大学资源学院,云南 昆明 650224)
采用径级结构代替年龄结构以及方差均值比率法对木兰科(Magnoliaceae)木莲属(ManglietiaBl.)濒危植物大果木莲(Manglietia grandisHu et Cheng)种群的年龄结构和种群格局进行了研究,并编制了大果木莲种群的特定时间生命表和存活曲线;结合生殖生物学特征以及遗传多样性研究结果,分析了导致大果木莲濒危的主要原因。根据株高和胸径可分别将大果木莲种群的年龄结构分为5级、高度结构分为6级;在大果木莲的5个年龄结构分级中,成年个体较多,幼年个体较少;其高度结构完整,个体高度主要在20 m以下。种群的方差均值比率为0.838 3,其空间分布格局属于随机分布。根据特定时间生命表可将大果木莲种群的发育分为3个阶段:幼树阶段(年龄级为Ⅱ~Ⅲ级)、成树阶段(年龄级为Ⅲ~Ⅳ级)、老树阶段(年龄级为Ⅳ~Ⅴ级),其中成树阶段个体死亡率最低。大果木莲种群存活曲线接近DeeveyⅠ型,属于衰退型种群。种群自我更新能力差、种子生产力低下、有性生殖困难、生境片断化导致的基因流受限以及人为干扰是大果木莲濒危的主要原因。针对大果木莲濒危现状和致危原因,提出了相应的保护对策和建议。
大果木莲;种群格局;年龄结构;生命表;存活曲线;濒危原因
Key words:Manglietia grandisHu et Cheng;population pattern;age structure;life table;survival curve;endangered cause
大果木莲(Manglietia grandisHu et Cheng)是国家二级重点保护野生植物及三级濒危保护物种,为木兰科(Magnoliaceae)木莲属(ManglietiaBl.)植物中分布比较狭窄的中国特有常绿乔木,零星分布于云南东南部和广西西南部[1-2]。大果木莲是木兰科中较为原始的类群,对研究古植物区系及木兰科分类系统和演化有重要的学术价值;其木材结构细致,耐腐、耐水湿、耐虫蛀,是建筑和家具的优良用材;其叶大而浓绿,花大而红艳、芳香,是中国南方十分重要的城市园林观赏树种[3-4]。然而,最近的调查结果表明:中国境内仅存1 200~1 500株大果木莲成熟个体,可能的分布范围约4 908 km2,而在滇东南的实际占有面积仅约0.3 km2[5],为此,大果木莲被世界自然保护联盟(IUCN)列为极度濒危植物(critically endangered species)[6]。因而,对大果木莲进行全面系统的濒危原因分析,并采取科学有效的保护措施已迫在眉睫。
关于大果木莲的地理分布、生物学和生态学、核型分析、种子休眠与萌发以及遗传多样性等方面皆有研究报道[3,7-11],但尚未见从种群生态学、生殖生物学及遗传多样性等角度对其濒危原因进行全面系统的研究报道。为此,作者根据种群格局特征分析了大果木莲的濒危原因,并提出相应的保护对策,旨在为大果木莲的有效保护提供理论指导和科学依据。
1 研究区概况和研究方法
1.1 研究区概况
样地位于云南大果木莲的主要分布区——文山州麻栗坡县,地理坐标为北纬22°48'~23°33'、东经104°33'~105°18',属南亚热带高原季风气候,以中低峡谷地貌为主,喀斯特地貌分布较广。大果木莲主要生长在海拔1 200~1 800 m沟谷或湿润山地的东南季风常绿阔叶林中。分布区气温较高,年平均气温17.5℃,1月(最冷月)平均气温10.1℃,7月(最热月)平均气温23℃;降水较多,年降雨量1 642~1 821 mm。伴生树种有硬斗石栎〔Lithocarpus hancei(Benth.)Rehd.〕、栲(Castanopsis fargesiiFranch.)、马关木莲(Manglietia maguanicaChang et B.L.Chen)、马关含笑(Michelia opiparaChang et B.L.Chen)、云南樟〔Cinnamomum glanduliferum(Wall.)Meisn.〕、木姜子(Litsea pungensHemsl.)、滇楠(Phoebe nanmuGamble)以及木荷(Schima superbaGardn.et Champ.)等;下木多为竹类;地被植物有秋海棠(Begonia evansianaAndr.)、麦冬〔Ophiopogon japonicus(L.f.) Ker-Gawl.〕等种类。土壤为黄壤和黄棕壤,pH 5.7~pH 6.5,自然含水率较高,有机质、全氮、有效氮、全磷、速效钾含量相对较高,有效磷和全钾含量较低。
1.2 研究方法
1.2.1 样方设置及调查在样地中设置面积10 m× 10 m样方43个,记录其中植物的种类以及大果木莲的株数、高度、胸径、盖度等指标。
1.2.2 年龄结构分级采用空间代替时间的方法,以立木级代替年龄结构[12],参照文献[13]采用经典分级方法、按照株高(H)和胸径(DBH)将大果木莲的年龄级分为Ⅰ级(H<33 cm)、Ⅱ级(H≥33 cm,DBH<2.5 cm)、Ⅲ级(2.5 cm≤DBH<7.5 cm)、Ⅳ级(7.5 cm≤DBH<22.5 cm)和Ⅴ级(DBH≥22.5 cm)。据此对样地内大果木莲个体进行年龄分级统计。
1.2.3 高度结构分级根据文献[14-15]的分级依据及实际调查结果,以株高(H)5 m为1个高度级差,将大果木莲分为Ⅰ级(H<5 m)、Ⅱ级(5 m≤H<10 m)、Ⅲ级(10 m≤H<15 m)、Ⅳ级(15 m≤H<20 m)、Ⅴ(20 m≤H<25 m)和Ⅵ级(25 m≤H<30 m) 6个高度级。据此统计样地内各高度级大果木莲的株数。
1.2.4 空间分布格局分析种群的空间分布格局用方差均值比率(S2/M)法判定[16]。即:在泊松分布中S2/M值等于或接近于1,为随机分布;S2/M值大于1,为集群分布;S2/M值小于1,为均匀分布。实测值与预期值的偏离程度采用t检验确定。
1.2.5 特定时间生命表编制用立木相对年龄的大小级代替绝对年龄级,根据特定时间生命表的编制原理和方法[17],计算生命表中的各项参数,编制大果木莲的特定时间生命表。
1.2.6 存活曲线绘制依据特定时间生命表,以立木的相对年龄大小级为横坐标,以In(lx)为纵坐标,绘制大果木莲的存活曲线。
2 结果和分析
2.1 大果木莲种群的年龄结构和高度结构分析
按照年龄级及高度级对43个样方中大果木莲的个体进行分级统计,结果见表1。在大果木莲的5个年龄结构分级中,Ⅰ级幼苗的数量为0,Ⅱ级幼树有29株,Ⅲ级、Ⅳ级和Ⅴ级的成树分别有31株、34株和5株。由此可以看出大果木莲种群中成年个体较多、幼年个体较少,属于衰退型种群。
由表1还可见:在Ⅰ至Ⅵ高度结构级中大果木莲的个体数分别为41株、30株、19株、7株、1株和1株。从高度级看,大果木莲种群的高度结构完整,种群中个体高度发育具有连续性,主要分布于20 m以下、特别是10 m以下的垂直空间中。
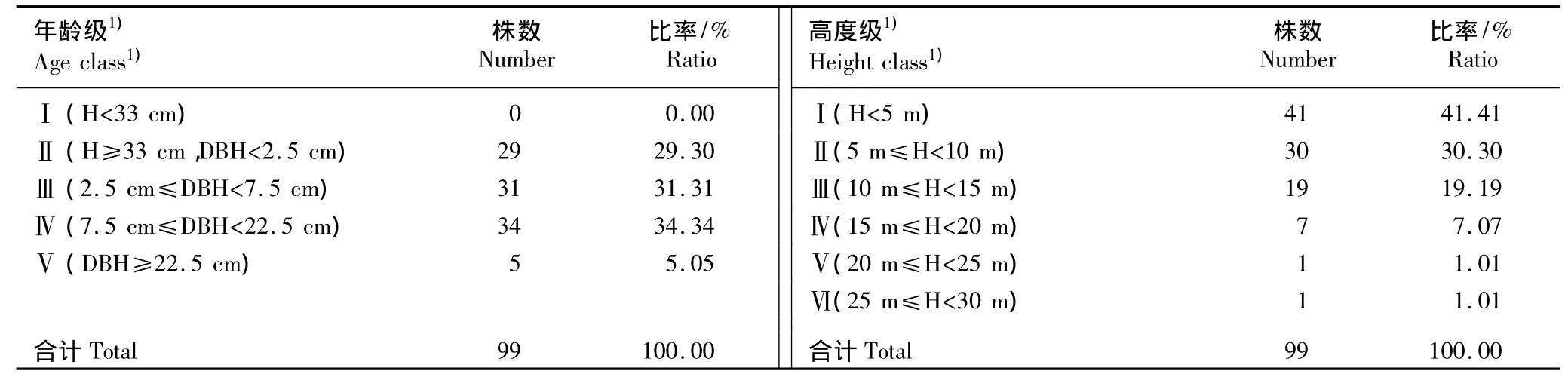
表1 大果木莲种群的年龄结构和高度结构分析Table 1 Analysis on age and height classes of Manglietia grandis Hu et Cheng population
2.2 大果木莲种群空间分布格局分析
大果木莲43个样方(n=43)在泊松分布中的方差S2=1.930 0、均值M=2.302 3,二者的比值S2/M= 0.838 3。用t检验检测实测值与预期值的偏离程度,t0.05(42)=2.019 0,大于∣t∣=0.741 0,S2/M接近1。按照泊松分布比值标准,大果木莲的空间分布格局属于随机分布。大果木莲在林分中是随遇种之一,在林下更新较差,成年植株周围很少有幼苗和幼树,呈现出随机分布的格局。
2.3 大果木莲种群的特定时间生命表
根据年龄结构分级数据,编制出大果木莲种群的特定时间生命表,结果见表2。由表2可见:大果木莲种群从幼苗生长到树木死亡大体可分为3个阶段:幼树阶段(年龄级为Ⅱ~Ⅲ级)、成树阶段(年龄级为Ⅲ~Ⅳ级)、老树阶段(年龄级为Ⅳ~Ⅴ级)。
幼树阶段个体死亡率较低,qx=-0.069。由于此阶段大果木莲个体间的竞争压力不大,因而只要温度和湿度等环境条件适宜,幼树大多能发育为成树。成树阶段个体死亡率最低,qx=-0.097。由于大果木莲大多分布于湿度较大且光照条件较好的林分内,因而这一时期植株均可获得充足的水分与光照以满足其生长所需,加之成树所具有的较强抗干扰能力,致使其死亡率最低。老树阶段个体死亡率最高,qx= 0.853。处于老年期的大果木莲植株生理功能衰退直至死亡,导致这一时期死亡率最高。

表2 大果木莲种群的特定时间生命表1)Table 2 The time-specific life table of Manglietia grandis Hu et Cheng population1)
2.4 大果木莲种群的存活曲线
依据特定时间生命表绘制大果木莲种群的存活曲线,结果见图1。根据Deevey对存活曲线的分型,大果木莲的存活曲线接近DeeveyⅠ型(即凸型)。根据Leak的划分标准[18],大果木莲种群属于衰退种群。

图1 大果木莲种群的存活曲线Fig.1 Survival curve of Manglietia grandis Hu et Cheng population
综合以上分析结果,大果木莲主要表现出以下3个种群生态学特征:地理分布及生境狭窄,种群数量极少;空间分布格局为随机型,属于衰退型种群;幼苗个体数少(无Ⅰ级幼苗),缺乏自我更新能力。
3 讨论和结论
3.1 大果木莲濒危原因分析
在1994年发布的全球IUCN红色名录中,大果木莲为“易受害”(VU)类植物,描述为:“分布区域减小(≤20 000 km2)、实际占有面积下降(≤2 000 km2)或变动大,被严重隔离或仅剩的生育地点数目≤10,且栖息地的范围、面积和品质持续下降、减少”。2003年,大果木莲成为濒危植物(EN),理由是:“据推测,过去3个世代内致危因素没有停止,种群至少减少50%”。由于占有面积持续降低及个体数量持续减少,目前该种被IUCN列为极度濒危植物(CR)。
大果木莲的濒危是多因素综合作用的结果,结合生殖生物学及遗传多样性的研究结果,导致大果木莲濒危的原因主要有以下几个方面:
3.1.1 种群极度缺乏自我更新能力大果木莲种群属于衰退类型,表现出濒危物种的特征,其根本原因是大果木莲种群几乎没有自我更新能力(主要表现为无Ⅰ级幼苗)。植物种群自我更新能力从某一方面直接反映其种群生存和发展的潜力,自我更新能力的丧失预示着种群的衰老和消亡。大果木莲种群自我更新极其困难的状况与其本身的生物学特性相关,例如:生殖受限或生殖所需的特殊条件得不到满足致使其再生困难、遗传多样性降低导致的适应力和生存力下降;同时也与外界因素密切相关,比如自我更新的条件和环境遭到自然或人为的严重干扰破坏,致使其无法实现自我更新。
3.1.2 种子生产力低下大果木莲具有较高的遗传多样性[10],说明大果木莲自我更新困难与遗传多样性低下导致的适应力和生存力降低无关。大果木莲繁育系统以异交为主,依靠昆虫传粉[11,19],野生状态下分布星散、开花时间差、花粉活性低且活性保持时间短,加之传粉昆虫及活动的制约,导致大果木莲实际种子生产力低下。另外,大果木莲种子败育较为严重,这也是导致种子生产力低下的原因之一。
3.1.3 有性生殖困难种子萌发直接影响植物的自然更新,而种子本身的特性及萌发条件又是决定种子萌发状况的两大主要因素。大果木莲种子具有“条件休眠”特性,属于浅度简单型形态生理休眠,可能还有休眠循环现象[5]。大果木莲新鲜种子萌发时间较长(约50 d),发芽持续时间很长(约11~17 d)[11],尽管平均发芽率不是很低(60%以上),但种子休眠和萌发特性影响了大果木莲在自然状态下的有性生殖效率。另外,大果木莲生境多为石灰岩山地,种子落地后不易接触到土壤,生境条件难以满足其种子萌发所需,致使大果木莲有性生殖困难。
3.1.4 生境片断化,基因流受限大果木莲种群间的遗传分化程度较高,基因流(Nm=0.890 8)较低[10],说明种群间的基因流动受到限制,强化了种群间的分化。大果木莲种群空间分布格局属于随机分布型,自然状态下星散分布于林分中,这样的分布格局及遗传结构格局暗示着大果木莲分布区片断化较为严重,对种群发展乃至物种生存都非常不利。
3.1.5 人类活动的影响人类活动的加剧、森林砍伐、城市化建设及土地资源的不合理利用所引起的生境丧失和片断化已经成为导致物种濒危的主要因素之一。人类活动对大果木莲的影响方式和程度是多种多样的,如:直接的采伐破坏、践踏和放牧对更新的破坏、毁林使生境干燥化渐变为“致危”生境等恶性影响。在众多的外因中,人为因子尤其是人类活动的强度和频度不断加剧,对大果木莲生存的影响是前所未有的。一方面,大果木莲是优良观赏树种,还是被称作“黄心木”的优良用材树种,其全株尤其树皮可作为“厚朴”药材提取厚朴酚等的原料之一,成为人们直接采挖和采伐的对象。另一方面,大果木莲生长于人类活动较频繁的山坡、路边,直接暴露于人类活动的影响之下,而且随着社会经济的发展,道路、工程建设和毁林开荒、乱砍滥伐、经济林种植等一系列高强度的人类活动破坏了大果木莲的适宜生境,造成其种群面积缩小且被分割成不同的生境斑块,形成破碎化的生境。大果木莲目前的种群结构及格局实际上主要就是人类干扰结果的体现。
3.2 大果木莲保护对策及建议
3.2.1 就地保护对大果木莲现有野生种群及其栖息地的保护是最为有效的保护措施。应尽可能将大果木莲分布区划入自然保护区范围内加强保护,尤其像遗传多样性较高的马关古林箐种群更应加强保护;而不能划入自然保护区的种群则应建立相应的保护站点加以保护,杜绝直接采挖、砍伐以及森林破坏、垦荒、放牧等活动。目前对分布于麻栗坡县铁厂乡和古林箐的大果木莲种群已采取就地保护措施,但对于分布于下金厂乡的大果木莲种群仍没有采取适当保护措施,砍伐和生境破坏等行为屡有发生。
3.2.2 迁地保育只要条件适宜大果木莲种子萌发率可高达70%,因而建议尽可能将大果木莲列为引种保育对象,建立大果木莲迁地保育基地。因其大部分的遗传变异(64.05%)存在于种群内[10],迁地保育时应尽可能收集各天然种群足够数量的单株,这样才能尽可能地保存大果木莲的遗传多样性,为归化自然、开发利用、科学研究等工作提供丰富的材料。
3.2.3 归化自然要获得濒危植物保护的完全成功,仅仅进行就地和迁地保护是不够的,还应该开展归化自然的工作,即将人工繁殖的大果木莲个体移植到其原有的生境中,让其归化至自然环境中,并正常生长发育、产生后代,扩大自然种群的规模。
3.2.4 扩大培育建立组织培养、扦插、嫁接等无性繁殖和人工林培育技术体系,通过开展大果木莲的人工规模化种植和产业化基地建设缓解对大果木莲自然资源的需求压力,进而达到保护自然资源的目的。
3.2.5 加强公众教育,提升保护意识野生植物的保护和管理是一项长期而艰难的工作,也是一项公众性的工作。因此,应通过科普宣传、普法活动和政策引导等途径加强公众生态道德和资源可持续利用的教育和宣传,对大果木莲的保护也有重要作用。
[1]郑万钧.中国树木志:第一卷[M].北京:中国林业出版社,1983:419-510.
[2]中国科学院中国植物志编辑委员会.中国植物志:第三十卷[M].北京:科学出版社,1996:82-198.
[3]张茂钦.云南珍稀濒危树种生态生物学研究[M].昆明:云南大学出版社,1998:204-216.
[4]李玉媛.云南国家重点保护野生植物[M].昆明:云南科学技术出版社,2005:199-244.
[5]潘睿,孙卫邦.极度濒危植物大果木莲种子的休眠与萌发[J].植物生理学通讯,2009,45(11):1089-1092.
[6]CICUZZA D,NEWTON A,OLDFIELD S.The Red List of Magnoliaceae[M].Cambridge:Lavenham Press,2007:26.
[7]龚洵,潘跃芝,杨志云.木兰科植物的杂交亲和性[J].云南植物研究,2001,23(3):339-344.
[8]孟爱平,王恒昌,李建强,等.中国木兰科11属40种植物的核形态研究[J].植物分类学报,2006,44(1):47-63.
[9]徐亮,司马永康,杨卫,等.大果木莲的花部数量变异研究[J].广西林业科学,2006,35(1):23-25.
[10]陈少瑜,韩燕,吴涛,等.木兰科濒危植物大果木莲遗传多样性的ISSR分析[J].福建林学院学报,2010,30(1):56-60.
[11]付玉嫔,陈少瑜,吴涛,等.中缅木莲与大果木莲种子形态特征及萌发特性的比较[J].西北林学院学报,2009,24(6): 33-37.
[12]张育新,马克明,祁建,等.北京东灵山海拔梯度上辽东栎种群结构和空间分布[J].生态学报,2009,29(6):2789-2796.
[13]王伯荪,李鸣光,彭少麟.植物种群学[M].广州:广东高等教育出版社,1995:94-98.
[14]康华靖,陈子林,刘鹏,等.大盘山自然保护区香果树种群结构与分布格局[J].生态学报,2007,27(1):389-396.
[15]戴月,薛跃规.濒危植物顶生金花茶的种群结构[J].生态学杂志,2008,27(1):1-7.
[16]李玉媛.菜阳河自然保护区定位监测[M].昆明:云南大学出版社,2003:97.
[17]李先琨,向悟生,唐润琴.濒危植物元宝山冷杉种群生命表分析[J].热带亚热带植物学报,2002,10(1):9-14.
[18]LEAK W B.Age distribution in virgin red spruce and Northern Hardwoods[J].Ecology,1975,56(5):1451-1454.
[19]付玉嫔,陈少瑜,吴涛.濒危植物大果木莲与中缅木莲的花部特征及繁育系统比较[J].东北林业大学学报,2010,38 (4):6-10.
(责任编辑:惠红)
Analyses on population pattern and endangered causes of endangered species Manglietia grandis Hu et Cheng
CHEN Shao-yu1,FU Yu-pin1,WU Tao1,SIMA Yong-kang1,HAO Jia-bo1,HAN Yan2(1.Yunnan Laboratory for Conservation of Rare,Endangered and Endemic Forest Plants of State Forestry Administration,Yunnan Key Laboratory for Forest Plant Cultivation and Utilization,Yunnan Academy of Forestry,Kunming 650204,China;2.Department of Resources,Southwest Forestry University,Kunming 650224,China),J.Plant Resour.&Environ.2012,21(2):102-106
Using the way of diameter structure instead of age structure and variance/mean ratio method,the age structure and population pattern of the endangered speciesManglietia grandisHu et Cheng belonging toManglietiaBl.in Magnoliaceae were studied,and the time-specific life table and survival curve of the species was created.And combining with research results of propagation and genetic diversity,the main endangered causes were analyzed.According to tree height and diameter of breath height,age structure ofM.grandispopulation is divided into five classes and its height structure into six classes.There are more adults and less young trees in five age classes.The height structure ofM.grandispopulation is complete and tree height is mainly below 20 m.The population pattern is characterized by a random distribution withS2/M=0.838 3.According to the time-specific life table,the development process ofM.grandispopulation can be divided into three stages:sapling stage(Ⅱ-Ⅲage class),mature tree stage(Ⅲ-Ⅳage class)and old tree stage(Ⅳ-Ⅴage class),with the lowest mortality rate in mature tree stage.The survival curve ofM.grandisis close to DeeveyⅠtype,meaningM.grandispopulation belongs to declined type.It is suggested that lacking regeneration,low seed production capability,sexual reproduction difficulty,limited gene flow caused by habitat fragmentation and intensive human activity are main endangered causes ofM.grandis.Aiming to endangered situation and causes ofM.grandis,the conservation strategies and proposals are put forward.
book=2012,ebook=50
Q948.12.1;X176
A
1674-7895(2012)02-0102-05
2011-08-31
云南省自然科学基金资助项目(2006C0066M)
陈少瑜(1968—),女,广东潮州人,博士,研究员,主要从事林木遗传与森林资源保护等方面的研究。
