人胚肺成纤维细胞复制性衰老的基因表达变化
2012-08-22李国栋童坦君张宗玉
李国栋 马 宏 童坦君 张宗玉
(北京大学基础医学院生化系,北京 100191)
环境和遗传是影响细胞和个体衰老进程的两大因素,尤其是遗传因素在衰老中发挥的作用正日渐得到学者们的重视。与肿瘤类似,衰老的启动与进展不是由单一基因操纵的,而是受到一系列基因的协同调节。早期研究发现衰老过程伴有一些基因表达的上调及另一些基因表达的下调,这些基因的变化可作为该组织或细胞衰老的特异性生物学标志。因此,寻找参与细胞或个体衰老过程的基因和基因调控网络是衰老基础研究的重要内容之一。基因芯片技术以核算杂交理论为基础,利用固定在芯片上的几千至几万条探针与样品的mRNA进行杂交,在一次实验中获取大量信息。基因芯片技术使用了光控固相化学、激光共聚焦等多项先进技术,实现了全部自动化,操作快速、简便。芯片技术的应用使得高通量分析细胞衰老等复杂基因调控网络成为可能。尽管近年利用芯片技术对组织以及细胞衰老的研究逐渐增多,但是针对人胚肺成纤维细胞(2BS)复制性衰老的基因筛查仍未见报道。在此,我们用含有4 096种人类基因的基因芯片,检测了2BS细胞在复制性衰老过程中的基因表达变化情况,结果显示衰老细胞与年轻细胞相比有117种基因表达的变化幅度在2倍以上,变化幅度在2.5倍以上的基因有46种,而且我们挑选的9种差异表达基因经RTPCR验证与芯片结果基本一致。
1 材料与方法
1.1 细胞培养 2BS细胞为衰老研究中心常规保存,细胞用含10%胎牛血清的DMEM培养基,于37℃、5%CO2条件下培养,细胞长至90%融合时,以1∶2分瓶传代。一般定义为:细胞代龄(PD)在PD30及以下的为年轻细胞,PD55及以上的为老年细胞。
1.2 基因芯片杂交 使用RNeasy mini kit(QIAGEN,Germany)试剂盒提取不同代龄2BS细胞的总RNA,细胞的收集和处理参照动物细胞的操作步骤。分光光度法对RNA定量并通过凝胶电泳对RNA完整性进行评价。mRNA的提取使用Oligotex mRNA Midi kit(QIAGEN,Germany)试剂盒。cDNA探针的制备和纯化参照许沈华等〔1〕方法进行,Cy3-dUTP和Cy5-dUTP分别用于标记年轻2BS细胞(PD28)和衰老2BS细胞(PD64)的cDNA。标记探针95℃变性5 min后,与含有4 096条人类全长基因的 HGEC-40S cDNA芯片 (Biostar Genechip,China)在60℃杂交15~17 h。杂交后的芯片用2×SSC+0.2%SDS、0.1×SSC+0.2%SDS、0.1%SSC 洗涤 3次,每次 10 min,室温晾干。检测结果使用GenePix 4000B扫描,数据分析使用Gene-Pix 3.0软件。通过预先选定40个管家基因作为内参基因对Cy3和Cy5的原始信号进行均衡和修正。同时为了监控芯片制备和杂交过程,设定水稻U2RNA、HCV外壳蛋白及空白液为阴性对照。
1.3 RT-PCR 从芯片结果中挑选9个在年轻和衰老2BS细胞中差异表达的基因进行RT-PCR验证,β-actin作为对照基因(各基因的引物序列和产物大小见表1)。PCR反应体系一般为20μl,包括 cDNA 模板3~5 μl,25 mmol/L MgCl21.6 μl,10 mmol/L dNTP 0.4μl,上下游引物的最终反应浓度为0.4μmol/L,1 U的Taq DNA聚合酶。首先95℃变性1 min,每次循环包括95℃变性20 s,56~60℃退火1 min,72℃延伸30 s,共30个循环。PCR产物经1%的琼脂糖凝胶电泳分离,溴化乙锭染色后,用凝胶成像系统进行扫描分析。
2 结果
2.1 年轻与衰老2BS细胞基因表达谱的差异分析 芯片检测结果中Cy5与Cy3信号强度的比值(Cy5∶Cy3)反映了年轻与衰老2BS细胞差异表达基因的相对水平,其中Cy5:Cy3大于2.0的基因定义为在衰老细胞中高表达的基因,而Cy5∶Cy3小于0.5的基因为在衰老细胞中低表达的基因。在检测的4 096条人类基因中,有117条基因的Cy5∶Cy3大于2.0或小于0.5,变化幅度在2.5倍以上的基因有46种,提示这些基因的表达很可能随着2BS细胞的衰老发生了变化,其中部分基因列于表2。基因功能归类分析显示,细胞衰老过程中下调的基因主要是细胞分裂或细胞周期相关基因,上调的基因则主要参与了细胞应激反应和蛋白分泌等生理过程,这与Ly等〔2〕报道的老年人皮肤成纤维细胞基因表达的变化相似。此外,差异表达的基因还涉及细胞骨架与细胞外基质重塑、物质代谢与蛋白修饰以及G蛋白偶联受体与离子通道等介导的信号转导过程(表2)。

表1 PCR扩增基因的引物序列及产物长度

表2 年轻2BS细胞与衰老2BS细胞基因表达谱的差异

续表2

续表2
2.2 RT-PCR验证部分基因在2BS细胞衰老过程中的变化我们挑选了9个在芯片检测中发生表达改变的基因(表1),它们在复制性衰老过程中的表达变化情况目前尚未见报道。分别提取 PD27、PD35、PD42、PD48、PD54、PD60、PD64 的 2BS 细胞总RNA并逆转录合成cDNA,用于对这些基因的RT-PCR验证和分析。在所检测的9个基因中,除Diubiquitin无明显变化外,其余8个基因的表达变化趋势与芯片检测结果一致,表达上调的基因有 Laminin α4、G0S8、GRK4、VCAM1 和 Selenoprotein P,下调的有Preproenkephalin、Mad2和TPKC1(图1)。
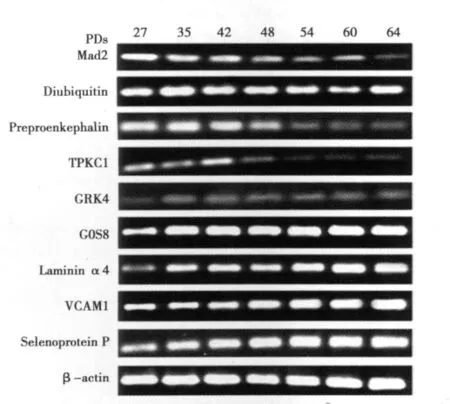
图1 RT-PCR检测部分差异表达基因在不同代龄2BS细胞的表达
3 讨论
体外培养细胞的复制性衰老作为一种个体衰老的简化模型,其可靠性已得到广泛的认可,然而基因调控网络的复杂性一定程度上阻碍了衰老分子生物学研究的进展。基因芯片技术是揭示衰老分子机制的一个重要手段,1999年Lee等〔3〕首次将基因芯片技术应用于衰老体系,发现限制热卡摄入可使老年鼠的基因表达谱向青年型转化。Lerner领导的科研小组应用基因芯片技术研究了老年人及Huchinson-giford早老症皮肤成纤维细胞基因表达情况,并将衰老推断为细胞分裂检控点失控的一种疾病〔2〕。在此,我们以含有4 096条人类基因的cDNA芯片检测了2BS细胞复制性衰老过程中的基因表达变化。cDNA芯片杂交结果显示:2BS细胞的复制性衰老并不引起基因表达的广泛变化,在所检测的4 096条人类基因中,荧光强度值变化在2倍以上的基因仅有117条。表达下调的基因主要参与细胞周期的进程,如在促有丝分裂因子MPF核转移过程中起作用的PLK和控制G2/M转换的Cyclin B。表达上调的基因则主要包括参与细胞应激反应和蛋白分泌等生理过程的分子,如凝血酶原活化因子(t-PA)、补体C3以及参与蛋白分泌途径的TRAMP分子。在差异表达基因中,有些基因已被证实在复制性衰老的成纤维细胞中高表达,如参与胎儿肺成熟的角质细胞生长因子(KGF)〔4〕随年龄增加在肺成纤维细胞中的表达明显升高,显示可能与成人伤口愈合情况比胎儿差有关〔5〕。
我们挑选并利用RT-PCR进一步检测了9个基因,它们在复制性衰老过程中的表达变化情况目前尚无报道,以下针对部分可能与衰老调节有关的基因进行讨论。Preproenkephalin是衰老细胞中下调幅度最大的基因,其在年轻2BS细胞中表达很强,从PD39左右表达开始下调,PD51以后表达极弱,但在过氧化氢诱导的早老细胞中未检测到前脑啡肽原基因表达的变化。研究显示胞核内脑啡肽水平的下降会导致抑郁行为,而前脑啡肽原敲除小鼠则不会出现抑郁表型〔6〕。鉴于Mianserin等抗抑郁药可能通过模拟节食的作用延长线虫的寿命〔7〕,因此前脑啡肽原及脑啡肽在细胞衰老中的作用值得进一步研究。衰老细胞下调幅度较大的基因中大部分与细胞有丝分裂进程相关,MAD2是细胞进入有丝分裂后期的检控点蛋白。cDNA芯片和RT-PCR检测结果均显示MAD2在2BS细胞复制性衰老过程中表达下调,而且我们在过氧化氢诱导的早老细胞中同样检测到MAD2 基因表达的下调〔8〕。
钾离子流动是膜电位改变的主要因素,有关神经突触和红细胞等衰老过程中伴有膜电位降低或钾离子流动性的改变已见诸文献〔9,10〕,但与成纤维细胞衰老的关系目前还未见报道。我们发现复制性衰老的2BS细胞中一种外向整流的钾离子通道TPKC1表达下调,而一种内向整流的TWIK-1的表达上调;RT-PCR也验证了TPKC1在2BS细胞衰老中的下调趋势。我们推测TPKC1表达的下调可能是衰老细胞膜电位降低的原因之一。G0S8又称G蛋白信号调节因子2(RGS2),是唯一选择性抑制Gqα的调节因子。2BS细胞衰老时G蛋白信号通路的负调控因子RGS2、RGS5及G蛋白耦联受体激酶GRK4表达上调。由于RGS2和GRK4的表达上调均发生在2BS细胞的青年或中年时期(图1),因此推测他们的表达变化可能是成纤维细胞复制性衰老的早期事件之一。此外,RGS2基因缺失的老年小鼠表现为体重较轻而且脂肪沉积减少〔11〕,提示可能参与调控生理代谢过程而影响衰老。硒蛋白是指含有硒半胱氨酸的一类蛋白质,人血浆中50%的硒元素是以硒蛋白P(Selenoprotein P)的形式存在的。近年研究表明:硒蛋白P可与过氧化亚硝酸及磷脂氢过氧化物反应,参与细胞外抗氧化防御机制〔12〕。芯片结果和RT-PCR检测均显示衰老2BS细胞中硒蛋白P表达上调,这与衰老细胞中DNA链断裂增加的现象似乎有些矛盾。Caldini等〔13〕曾报道:随体外培养的MRC5成纤维细胞倍增次数的增加,细胞内另外一种含硒的、参与清除自由基的蛋白质-谷胱甘肽过氧化物酶活性增强。因此我们推测衰老2BS细胞中硒蛋白P基因表达的上调是细胞复制性衰老过程中细胞内外环境活性氧基团浓度升高导致的诱导性调节。Miles等〔14〕发现人体血浆中血管细胞黏附分子1(VCAM1)的浓度与年龄有良好的正线形关系,这与我们的检测结果具有相同的变化趋势;然而Shelton等〔15〕观察到VCAM1在人脐带血管内皮细胞体外培养传代过程中显著下调。研究表明:转入端粒酶催化亚单位基因可使内皮细胞与成纤维细胞永生化,而且这两种细胞复制性衰老时均表现端粒缩短、衰老相关β-半乳糖苷酶染色增加及特定的细胞周期分布,这说明体外培养内皮细胞与成纤维细胞经历相似的衰老启动事件,VCAM1表达差异与细胞的特异性有关;而且它的表达变化不是启动复制性衰老的早期事件,而是细胞衰老的效应分子。
综上所述,2BS细胞复制性衰老基因表达变化的主要特征是细胞周期进程相关基因的下调和细胞应激反应及分泌蛋白类基因的上调。基因表达谱的分析是细胞衰老和个体衰老研究中的一个重要手段,然而由于通过芯片等高通量分析技术获得的基因信息往往数量巨大,难以建立简单易测的分子生物学衰老指征。如果在对基因信息进行生物学分析的基础上,针对部分感兴趣的基因或联合其他细胞学指征进行统计学分析将有助于细胞衰老的定性和定量评价,也有助于对细胞衰老基因调控网络的研究。
1 许沈华,牟瀚舟,吕桂泉,等.高低转移人卵巢癌细胞系基因表达谱差异〔J〕.中国肿瘤,2001;10(1):41-3.
2 Ly DH,Lockhart DJ,Lerner RA,et al.Mitotic misregulation and human aging〔J〕.Science,2000;287:2486-92.
3 Lee CK,Klopp RG,Weindruch R,et al.Gene expression profile of aging and its retardation by caloric restriction〔J〕.Science,1999;85:1390-3.
4 Shannon JM,Pan T,Nielsen LD,et al.Lung fibroblasts improve differentiation of rat type Ⅱ cells in primary culture〔J〕.Am J Respir Cell Mol Biol,2001;24(3):235-44.
5 Maas-Szabowski N,Shimotoyodome A,Fusenig NE.Keratinocyte growth regulation in fibroblast cocultures via a double paracrine mechanism〔J〕.J Cell Sci,1999;112(12):1843-53.
6 Andras B,Kerstin M,Florence N,et al.Preproenkephalin knockout mice show no depression-related phenotype〔J〕.Neuropsychopharmacology,2007;32:2330-7.
7 Petrascheck M,Ye X,Buck LB.An antidepressant that extends lifespan in adult Caenorhabditis elegans〔J〕.Nature,2007;450(7169):553-6.
8 马 宏,张 宗,童坦君.H2O2诱发人成纤维细胞衰老样变化的基因表达谱〔J〕.生物化学与生物物理进展,2003;30(1):72-7.
9 Ross ST,Soltesz I.Selective depolarization of interneurons in the early posttraumatic dentate gyrus:involvement of the Na(+)/K(+)-ATPase〔J〕.JNeurophysiol,2000;83(5):2916-30.
10 Reiff DF,Guenther E.Developmental changes in voltage-activated potassium currents of rat retinal ganglion cells〔J〕.Neuroscience,1999;92(3):1103-17.
11 Nunn C,Zhao P,Zou MX,et al.Resistance to age-related,normal body weight gain in RGS2 deficient mice〔J〕.Cell Signal,2011;23(8):1375-86.
12 Mostert V.Selenoprotein P:properties,functions,and regulation〔J〕.Arch Biochem Biophys,2000;376(2):433-8.
13 Caldini R,Chevanne M,Mocali A,et al.Premature induction of aging in sublethally H2O2-treated young MRC5 fibroblasts correlates with increased glutathione peroxidase levels and resistance to DNA breakage V〔J〕.Mech Ageing Dev,1998;105(1-2):137-50.
14 Miles EA,Thies F,Wallace FA,et al.Influence of age and dietary fish oil on plasma soluble adhesion molecule concentrations〔J〕.Clin Sci(Lond),2001;100(1):91-100.
15 Shelton DN,Edwin C,Whittier PS,et al.Microarray analysis of replicative senescence〔J〕.Curr Biol,1999;9:939-45.
