亚洲带绦虫六钩蚴Ta 8基因的克隆表达及免疫原性分析*
2012-08-21程金芝戴佳琳陈小睿廖兴江
王 宇,程金芝,黄 江,戴佳琳,陈小睿,廖兴江
2.贵州省疾病预防控制中心,贵阳 550004
至20世纪70年代以来,在我国台湾地区、东南亚地区和我国大陆地区相继报道了一种带绦虫,其成虫特征几乎和牛带绦虫一致,而囊尾蚴又与猪带绦虫极为相似。这种带绦虫的中间宿主是猪,人因生食猪肝等内脏而感染。这种带绦虫被命名为亚洲带绦虫[1]。近年来的研究证实在贵州省都匀地区存在亚洲带绦虫病的流行[2]。但是由于传统的病原学方法诊断该病较困难,尤其易与牛带绦虫混淆,使免疫学方法以其费用低廉、操作简单、特异性高等特点成为目前诊断中的重要辅助诊断手段,其中筛选特异性好、敏感性高的诊断抗原成为免疫学诊断技术研究的关键。本研究通过构建p ET-28a(+)-Ta8原核表达载体,并对其原核表达产物进行纯化和免疫学初步研究。筛选具有免疫原性和潜在诊断价值的候选抗原基因,为绦、囊虫病免疫诊断方法的建立奠定基础。
1 材料与方法
1.1 材料
1.1.1 标本来源,质粒,菌株 虫体标本采自亚洲带绦虫流行区贵州省都匀市米秀乡;原核表达质粒p ET-28a(+)和大肠杆菌BL21/DE3由中山大学热带病重点实验室惠赠。
1.1.2 主要试剂和工具酶 TRIzol试剂购自美国Invitrogen公司,逆转录试剂盒(PrimeScriptTMRT reagent Kit)购自日本Ta KaRa公司,Ex Taq酶、EcoR I、HindⅢ、DNA标准(DL15 000、DL2 000)均购自大连宝生物工程公司,T4 DNA连接酶、异丙基-β-D-硫代半乳糖苷(IPTG)均购自美国 Promega公司,Ni-IDA Agarose(Cat.No.30210)购自德国NOVEGN公司,兔抗鼠Ig G试剂盒购自武汉博士德生物工程有限公司,质粒提取试剂盒购自广州东盛科技公司,低分子量蛋白分子量标准购自立陶宛Fermentas公司,DNA凝胶回收试剂盒购自广州铂尔生物科技公司。
1.1.3 引物的合成和DNA的测序 基因扩增引物由Invitrogen上海生物技术有限公司合成,重组质粒DNA的测序由金斯特科技股份有限公司完成。
1.2 方法
1.2.1 亚洲带绦虫虫卵孵化、六钩蚴总RNA的提取及c DNA的合成 取新鲜亚洲带绦虫成虫孕节,用灭菌生理盐水清洗干净,用剪刀剪碎,4 000 r/min离心5 min。用灭菌生理盐水洗3次(4 000 r/min离心5 min),用显微镜计数。将分离的虫卵放入培养皿中,加入次氯酸钠溶液,使虫卵胚膜破裂,待胚膜破裂完后立即用生理盐水洗3次,沉淀用少量生理盐水重悬沉淀,加入人工胰液,37℃水浴40 min激活六钩蚴,用生理盐水洗3次后,将沉淀立刻放进DEPC水处理的匀浆器中,加入1 m L TRIzol后匀浆,直至样品全部融化成液体,将其移入无菌且RNase free EP管,用力振荡3 min,室温静置5 min。加入0.2 m L氯仿,混合后静置10 min,4℃12 000 g离心20 min,小心取上清液到一个新的RNase free EP管中。加入0.5 m L异丙醇,室温静置10 min,4℃12 000 g离心15 min,弃上清液。沉淀加入1 m L 75%乙醇,震荡15 s,4℃7 500 g离心5 min,弃上清液,倒置在超净台上10 min,加入30 μL纯水溶解沉淀,所获得的总RNA保存于-80℃冰箱中。取适量的RNA样品合成cDNA,反应体系为:模板2μL(500ng),5×PrimeScript缓冲液2 μL,PrimeScript酶混合物0.5μL,Oligod T和Random 6 mers引物各0.5μL,加RNase Free d H2O至总体系为10μL。反应条件:37℃30 min,85℃5 s。-20℃保存备用。
1.2.2 引物的设计合成、基因扩增 根据Gen-Bank上猪带绦虫相关8 k Da基因序列,用Premier 5.0 设 计 引 物,P1:5′-CGCGAATTCGTATCGGCTTGTAAAATT-3′,带EcoR I酶切位点;P2:5′-CGCAAGCTTTTAAGCAGTTTTGTTCTTCAGAC-3′,带HindⅢ酶切位点。以亚洲带绦虫六钩蚴cDNA为模板,应用上述设计的特异引物扩增目的基因,PCR产物行1.5%琼脂糖凝胶电泳鉴定回收、测序鉴定。
1.2.3 检测Ta8基因在亚洲带绦虫中不同时段、以及成虫不同部位的表达情况 成虫头节、成节、孕节总RNA的提取及cDNA的合成除不用孵化虫卵外其余步骤与提取六钩蚴总RNA相同。采用RTPCR方法用合成的特异引物分别扩增六钩蚴、成虫头节、成节、孕节总RNA中的Ta8基因,行1.5%琼脂糖凝胶电泳检测结果。
1.2.4 重组原核表达质粒的构建及鉴定 将六钩蚴PCR产物胶回收后和原核表达质粒p ET-28a(+)用EcoRI和HindⅢ进行双酶切后回收,连接、转化大肠埃希菌BL21/DE3感受态细胞,卡拉霉素筛选阳性单克隆,提取的重组质粒进行双酶切、PCR和DNA测序鉴定后进行原核表达。
1.2.5 重组蛋白的诱导表达 挑取含有p ET-28a(+)Ta8重组质粒的E.coliBL21/DE3的单菌落接种于含卡那霉素的LB液体培养基中(空质粒p ET-28a(+)的BL21/DE3作为阴性对照),放入37°C摇床,摇至OD600约为0.6时,取1 m L作为诱导前样品,加入IPTG至终浓度为1 mmol/L,摇床诱导表达5 h后离心收集菌体作为诱导后样品,所有样品离心后收集菌体处理后离心取上清行SDS-PAGE电泳分析。
1.2.6 重组蛋白的纯化、抗体制备和蛋白质印迹(Western blot)分析 依据上述诱导表达方法对阳性克隆进行大量的诱导表达,离心收集菌体,冰上超声裂解,分别取上清和沉淀样品处理后行SDSPAGE判断重组蛋白的可溶性。尿素变性纯化重组蛋白,按参考文献[3]方法进行,用Bradford法测蛋白浓度,并于-80℃保存备用。用纯化的蛋白与弗氏佐剂混匀免疫SD大鼠,每2 w免疫1次,第3次免疫后1 w取大鼠血清分离抗体,小份分装于-20℃保存备用,用前灭活。纯化的Ta8蛋白经SDS-PAGE电泳后转移至硝酸纤维素膜(NC膜),用5%脱脂奶粉封闭过夜,第2 d取出后用PBS洗涤3次,用NC膜与健康大鼠血清和Ta8蛋白抗原免疫大鼠血清(作为一抗,1∶100)室温孵育2 h,用PBS洗涤3次,5 min/次。再将膜与兔抗鼠HRPIgG(作为二抗,1∶2 000)室温孵育1.5 h,PBS洗涤3次,加入二氨基联苯胺(DAB)试剂显色,观察结果。
2 结 果
2.1 RNA提取和8 k Da序列分析 对亚洲带绦虫六钩蚴提取总RNA,紫外分光光度计测定OD260/280为2.02,取部分 RNA合成cDNA,RTPCR扩增目的基因后胶回收,通过测序后用生物信息学对序列进行识别分析。从BLASTx的分析结果来看,该基因与猪带绦虫8 k Da基因(登录号为AAM00212.1)的氨基酸序列的同源性达80%,所以推测为亚洲带绦虫六钩蚴8 k Da基因,含有一个最长的ORF就是其完整的编码区,起始密码为ATG,终止密码为TAA,编码83个氨基酸,见图1。其理论分子量和等电点分别为9 416.1 Da和9.47。

图1 Ta 8基因序列及其ORF编码的氨基酸序列Fig.1 Sequence of Ta 8 gene and the amino acid sequence encoded by the ORF
2.2 检测Ta8基因在亚洲带绦虫中不同时段、以及成虫不同部位的表达情况 采用RT-PCR方法,通过特异性扩增六钩蚴、成虫头节、成节、孕节cDNA中的Ta8基因,结果显示Ta8基因不仅在六钩蚴中表达,在成虫的不同部位也同时表达,见图2。

图2 Ta 8基因在六钩蚴、成虫头节、成节、孕节阶段的表达1:六钩蚴;2:头节;3:成节;4:孕节;M:DNA 标志物;5:阴性对照Fig.2 The expression of Ta 8 gene in oncosphere,scolex,and mature and gravid proglottids1:Oncosphere;2:Scolex;3:Mature proglottids;4:Gravid proglottids;M:DNA marker;5:Negative control
2.3 原核重组质粒的鉴定、分析 将重组质粒进行PCR和双酶切鉴定,产物行1.5%琼脂糖凝胶电泳。结果显示约240 bp处有一清晰的条带,与目的基因的大小基本相符,见图3,测序证明重组质粒构建成功。
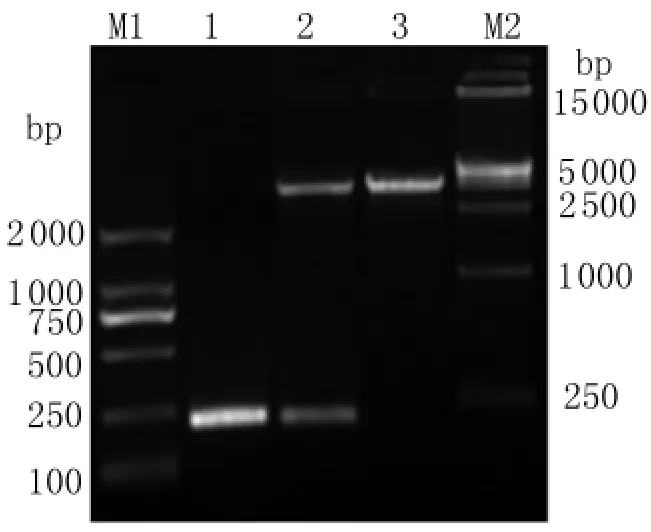
图3 重组质粒的PCR及双酶切鉴定M1:DNA标志物2 000;1:从亚洲带绦虫六钩蚴cDNA中扩增的 PCR 产物;2:重组质粒(p ET-28a(+)-Ta8)用Eco RⅠ和 Hin dⅢ酶切;3:重组质粒(p ET-28a(+)-Ta 8);M2:DNA标志物15 000。Fig.3 Identification of the pET-28a(+)-Ta8 by PCR amplification and digestion with restriction enzymesM1:DNA marker DL2000;1:PCR product amplified from cDNA of Taenia asiatica oncosphere;2:Recombinants(p ET-28a(+)-Ta8)digested by Eco RⅠ and Hin dⅢ;3:Recombinants(p ET-28a(+)-Ta8);M2:DNA marker DL15000
2.4 蛋白表达纯化结果 将构建好的重组质粒转化到E.coliBL21/DE3中表达,SDS-PAGE电泳分析结果第4、6泳道所示,大约在11 k Da左右处出现表达条带,与目的蛋白分子量基本相符。将蛋白进行纯化,结果第7泳道所示,证明目的蛋白纯化成功。用Bradford法测该蛋白浓度为0.65 mg/m L,见图4。
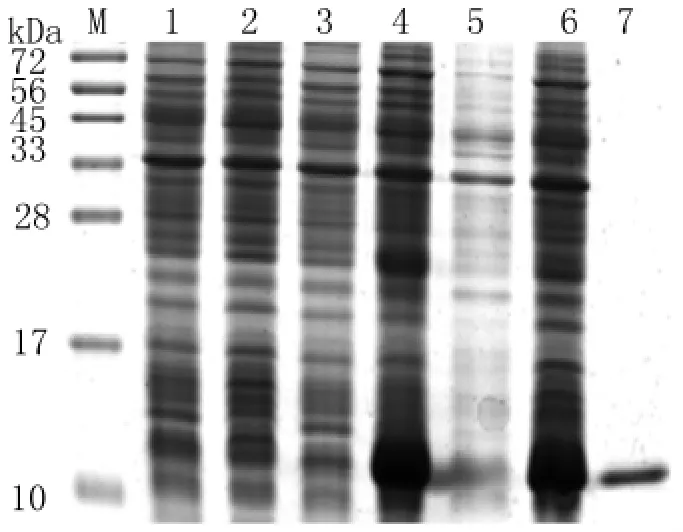
图4 重组质粒的表达及其纯化产物的15%SDS-PAGE电泳分析M:蛋白标志物;1:p ET-28a(+)质粒未加IPTG 诱导;2:p ET-28a(+)质 粒 加 IPTG 诱 导;3:p ET-28a(+)-Ta 8质粒未加IPTG 诱导;4:p ET-28a(+)-Ta 8质粒 加IPTG 诱 导;5:p ET-28a(+)-Ta 8质 粒 加IPTG诱 导 后 上 清;6:p ET-28a(+ )-Ta 8 质 粒 加IPTG诱 导 后 沉 淀;7:p ET-28a(+ )-Ta 8 质 粒 加IPTG诱导后纯化产物。Fig.4 15%SDS-PAGE analysis of the prokaryotic expression product and purified productM:Protein marker;1:p ET-28a(+)transformants without IPTG induction;2:p ET-28a(+)transformants with IPTG induction;3:p ET-28a(+)-Ta 8 transformants without IPTG induction;4:p ET-28a(+ )-Ta 8 transformants with IPTG induction;5:Insoluble protein of p ET-28a(+)-Ta 8 transformants;6:Sediments of lysate p ET-28a(+)-Ta 8 with IPTG induction;7:Purification of insoluble protein of p ET-28a(+)-Ta 8 transformants
2.5 Western blot鉴定重组蛋白Ta8的免疫原性
纯化得到的重组蛋白Ta8行15%SDS-PAGE,Western Blot分析,使用SD大鼠抗重组蛋白Ta8的特异抗血清作为一抗,结果显示特异性抗血清能识别重组蛋白Ta8,而正常大鼠血清不能识别,见图5。
3 讨 论
亚洲带绦虫与牛带绦虫的形态结构很相似,使用传统的病原学方法诊断该病较困难,目前对亚洲带绦虫的诊断最可靠的方法是靠驱虫后对该虫线粒体Cox1基因片段的鉴定和形态结构特征共同确定虫种[4],但是该技术操作繁琐,使免疫学方法以其费用低廉、操作简单、特异性高等特点成为目前诊断中的重要辅助诊断手段,其中筛选特异性好、敏感性高的诊断抗原成为免疫学诊断技术研究的关键。

图5 重组Ta 8蛋白的免疫原性M:蛋白标志物;1:重组蛋白免疫SD大鼠后血清;2:健康大鼠血清。Fig.5 Immunogenicity of recombinant Ta 8M:Protein marker;1:Recombinant p ET-28a(+)-Ta 8 reacted with serum from immunized SD rat;2:Crude antigens reacted with serum from normal rat
目前,用于带绦虫科诊断研究的抗原不多,8 k Da蛋白为其中之一。有研究报道带绦虫科8 k Da蛋白具有很高的免疫原性和诊断价值,可作为带绦虫病或者囊虫病的诊断抗原[5-6]。带绦虫科8 k Da蛋白是一个很大的家族,每个基因之间序列差别较大,Hancock等[7]表达了猪带绦虫18种8 k Da蛋白,其中9种8 k Da蛋白经ELISA检测,发现对人阳性猪囊虫病血清表现出的敏感性和特异性都为100%,因此可以推测亚洲带绦虫8 k Da蛋白也可能是亚洲带绦虫病的重要免疫诊断抗原之一。
成功从亚洲带绦虫六钩蚴的cDNA中获得8 k Da基因,并构建了p ET-28a(+)-Ta8重组质粒,测序后序列经NCBI网站Blastx在线分析,该基因与猪带绦虫8 k Da基因(登录号为AAM00212.1)的氨基酸序列的同源性达80%,含有一个最长的ORF就是其完整的编码区,编码83个氨基酸,其理论分子量和等电点分别为9 416.1Da和9.47。蛋白质的理化性质较稳定。Western Blot结果表明重组蛋白能被其免疫的SD大鼠血清识别而不能够被正常SD大鼠血清识别,说明重组蛋白具有免疫原性。采用RT-PCR方法检测8 k Da基因在亚洲带绦虫六钩蚴、以及成虫不同部位的表达情况,发现8 k Da基因不仅在六钩蚴阶段表达,在成虫头节、成节、孕节中均表达,这样就大大提高了采用血清来诊断亚洲带绦虫病的可能性,为进一步开展诊断试剂盒或试纸条的研究提供了有利条件。
综上所述,8 k Da基因是一个庞大的家族,要全面的了解其成员的功能,必须进一步研究其同源的基因,才能更好地筛选潜在的候选抗原。
[1]Fan PC,Lin CY,Chung WC.Experimental infection of PhilippineTaeniain domestic animals[J].Int J Parasitol,1992,22(2):235-238.DOI:10.1016/0020-7519(92)90107-V
[2]Bao HE.The current situation and prospect ofTaeniasaginata asiaticain China[J].J Trop Med,2002,2(3):215-219.(in Chinese)
包怀恩.我国亚洲牛带绦虫研究的现状和展望[J].热带医学杂志,2002,2(3):215-219.
[3]Shen PX,Wu X,Huang J,et al.Prokaryotic expression of elongation factor 1 gene ofTaeniasaginataasiaticaand analysis of purification and immunogenicity of the recombinant protein[J].J Xi'an Jiaotong Univ(Med Sci),2008,29(4):379-382.(in Chinese)
申萍香,吴 璇,黄 江,等.亚洲牛带绦虫成虫延伸因子-1基因的表达、纯化及免疫学分析[J].西安交通大学学报,2008,29(4):379-382.
[4]Chen ZH,Bao HE,Mou R,et al.Molecular differentiation of 38 human taeniid cestodes from Yunnan and Guizhou provinces[J].Chin J Pathog Biol,2010,5(4):263-265.(in Chinese)
陈峥宏,包怀恩,牟荣,等.云南、贵州两省38条人体带绦虫的分子鉴别[J].中国病原生物学杂志,2010,5(4):263-265.
[5]Jiang L,Xue HC,Qiu LS,et al.Western blotting analysis of specific antigens from different components ofEchinococcus metacestodes[J].Chin J Parasitol Parasit Dis,2004,22(1):11-15.(in Chinese)
江莉,薛海筹,裘丽姝,等.棘球蚴特异性抗原的蛋白质印迹分析[J].中国寄生虫学与寄生虫病杂志,2004,22(1):11-15.
[6]Haag KL,Zanotto PM,Alves-Junior L,et al.Searching for antigen B genes and their adaptive sites in distinct strains and species of the helminthEchinococcus[J].Infect Genet Evo,2006,6(4):251-261.DOI:10.1016/j.meegid.2005.07.003
[7]Hancock K,Khan A,Williams FB,et al.Characterization of the 8-kilodalton antigens ofTaeniasoliummetacestodes and evaluation of their use in an enzyme-linked immunosorbent assay for serodiagnosis[J].J Clin Microbiol,2003,41(6):2577-2586.DOI:10.1128/JCM.41.6.2577-2586.2003
