3种生态型野生扁蓿豆种质资源ISSR与SSR遗传多样性分析
2012-08-20李鸿雁李志勇师文贵蔡丽艳张静萍
李鸿雁,李志勇*,师文贵,蔡丽艳,张静萍
(1.中国农业科学院草原研究所 农业部沙尔沁牧草资源重点野外科学观测试验站,内蒙古 呼和浩特010010;2.内蒙古大学交通学院,内蒙古 呼和浩特010021)
苜蓿属扁蓿豆植物在内蒙古境内有1个正种、3个变种,即扁蓿豆(Medicago ruthenica)、细叶扁蓿豆(M.ruthenicavar.oblongifolia)、辽西扁蓿豆(M.ruthenica var.liaosiensis)和阴山扁蓿豆(M.ruthenica var.inschanica)[1]。从形态分类上目前有学者在内蒙古发现扁蓿豆新类型,即野生黄花型扁蓿豆[1]。扁蓿豆是苜蓿遗传改良的重要优异基因来源,具有重要的利用价值。近年来国家牧草种质中期库收集、保存、鉴定评价了210份野生扁蓿豆种质资源,对筛选出的优良材料从形态学水平、细胞水平和DNA水平进行遗传多样性的系统研究。
SSR和ISSR等技术已先后应用于该植物的遗传多样性研究[2-6],基于DNA-PCR的分子标记技术已成为形态学之外的主要分析手段之一。不同分子标记在遗传多样性研究中表现出各自的特性。选择合适的分子标记对能否客观反映研究对象的状况具有重要影响。ISSR(inter-simple sequence repeat)是一种利用重复序列加选择性碱基为引物,对基因组DNA进行扩增的标记[6],ISSR引物序列长,退火温度高,特异性和稳定性增强,可检测到更丰富的遗传变异。ISSR技术在一些重要的牧草,如无芒雀麦(Bromus inermis)、昆仑锦鸡儿(Caragana polourensi)、小花棘豆(Oxytropis glabra)和新麦草(Psathyrostachys juncea)的遗传多样性分析方面取得了很大进展[7-10]。Martin和Sanchez-Yelamo[11]研究表明ISSR标记与形态学、生化及其他分子标记存在较高一致性。SSR(simple sequence repeat)是目前应用最为广泛的分子标记技术之一,具有理想分子标记绝大多数的优良特点,且苜蓿属SSR引物在种属间具有一定通用性[12],近年来,微卫星技术广泛应用于披碱草(Elymus dahuricus)、鸭茅(Dactylis glomerata)、苜蓿(Medicago sativa)、锦鸡儿属(Caragana)等牧草的遗传多样性分析中[13-16]。本研究选取二倍体的黄花型扁蓿豆、细叶扁蓿豆和扁蓿豆为实验材料,对ISSR和SSR标记在扁蓿豆种质资源研究的可应用性进行探讨,并结合种间关系分析比较两者的标记效率和多样性检测能力。
1 材料与方法
1.1 材料
供试材料为6份黄花型扁蓿豆、2份细叶扁蓿豆和6份扁蓿豆,均为国家牧草中期库课题组野外考察收集的材料,原产内蒙古(表1)。于2006年种植在农业部沙尔沁牧草资源重点野外科学观测试验站。经田间形态鉴定评价后,分别于2008年和2009年在中国农业科学院国家农作物种质资源保存中心实验室及国家牧草种质中期库分子生物学实验室进行SSR和ISSR标记分析。
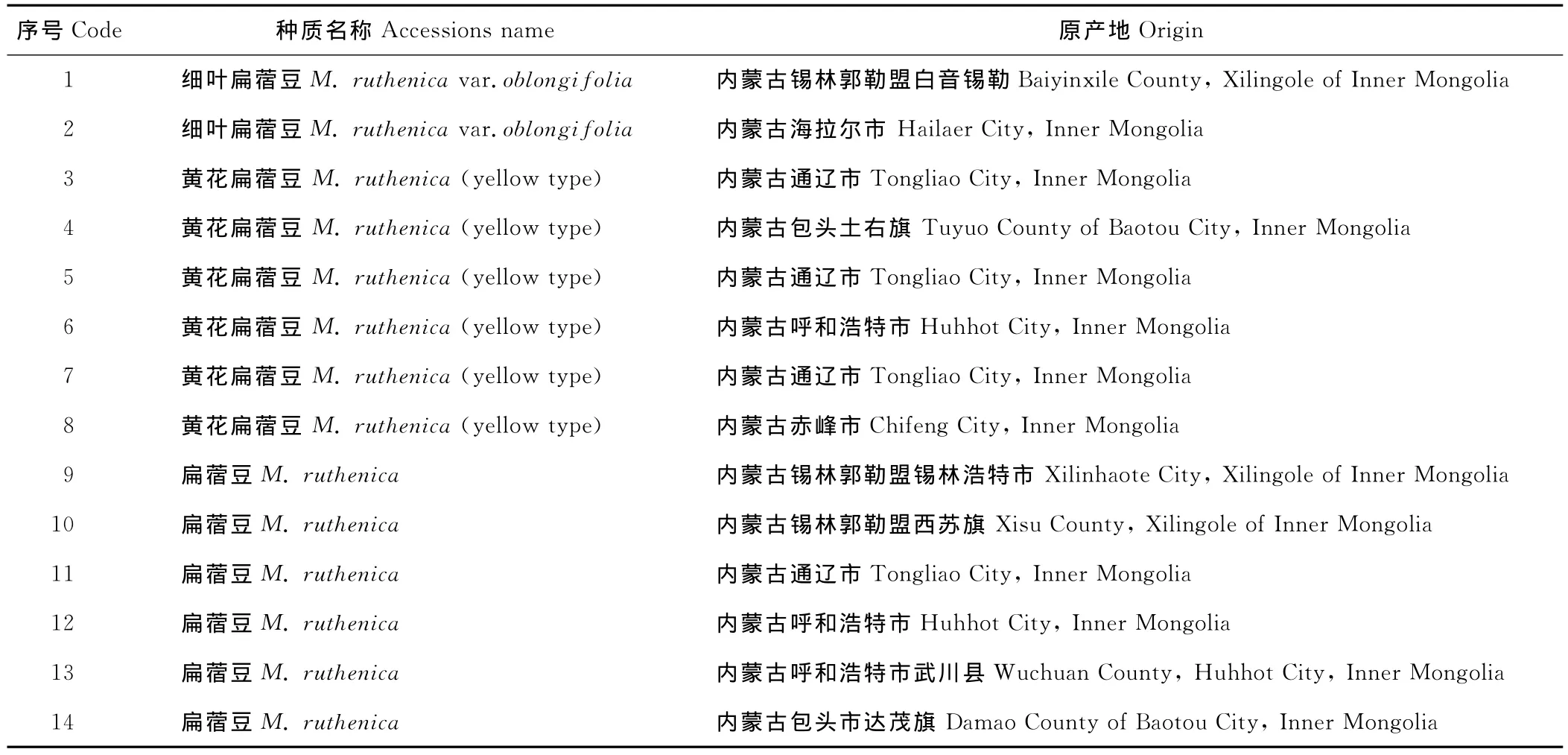
表1 14份扁蓿豆种质来源及编号Table 1 The code and origin of the 14accessions of M.ruthenica
1.2 PCR扩增
1.2.1 DNA提取 每份材料随机选取30个单株的新鲜叶片,用SDS法略加改进提取每个单株叶片基因组DNA。
1.2.2 ISSR扩增和检测 参考加拿大哥伦比亚大学提供的二核苷酸重复ISSR引物中筛选出多态性丰富的18个引物(表2),对14份材料420个样品进行PCR扩增。25μL PCR反应体系:模板DNA 0.5ng/μL,10×buffer(含 Mg2+)2.0mmol/L,TaqDNA聚合酶1.5U,dNTPs 0.6mmol/L,引物0.9μmol/L。PCR扩增:扩增程序为94℃预变性3min,94℃变性30s,50℃复性45s,72℃延伸1.5min,循环35次;72℃延伸10min,4℃保存。用1.5%琼脂糖凝胶电泳检测扩增产物,电极缓冲液为1×TAE,电泳时间40min,用DL2000作对照。电泳结束后观察照相。ISSR引物、dNTPs及TaqDNA聚合酶均购自上海生物工程技术服务有限公司。
1.2.3 SSR扩增和检测 根据Bernadette等[17]的报道,从89对紫花苜蓿和截形苜蓿SSR引物中筛选多态性丰富、重复性好的18对引物进行扁蓿豆PCR扩增(表2)。PCR反应体系为25μL:模板DNA 3μL,10×Buffer(含 Mg2+)5.5μL,Taq DNA聚合酶(由北京天根生物公司合成)0.5μL,dNTP 0.75μL,引物(10mmol/L)0.75μL,ddH2O 14.5μL。PCR反应程序为:95℃预变性3min;94℃变性25s,55~66℃(根据不同引物而定)退火30s,72℃延伸30s,35个循环;72℃延伸10min,最后4℃保存。取扩增产物,经6%变性聚丙烯酰胺凝胶电泳检测(恒功率80W,30min)后,进行银染、固定、染色和显影。引物由上海生物工程技术服务有限公司合成。
1.3 数据分析
以清晰可辨的扩增条带在相对迁移位置的有无记数,有扩增带时,赋值为“1”,无扩增带时赋值为“0”,生成分子数据矩阵。利用Popgen 3.2统计SSR数据和ISSR数据的遗传相似系数(GS),Shannon指数、Nei’s指数,利用 NTSYSpc 2.1软件[18]采用 UPGMA (Unweighted Pair Group Method with Arithmetic mean)法进行聚类分析,绘制亲缘关系树状聚类图,用Mantel检验进行相关性检测。

表2 ISSR和SSR的引物序列Table 2 The prime sequences of ISSR and SSR
2 结果与分析
2.1 扩增结果的多态性分析
18个ISSR引物可扩增出清晰而重复性好的条带,且具多态性,18对紫花苜蓿和截形苜蓿SSR引物可扩增出清晰谱带,对14份扁蓿豆材料进行PCR扩增。
14份扁蓿豆的ISSR扩增结果见图1和表3,ISSR扩增出的条带共143条,条带的分子量范围为250~2 000 bp,其中多态性条带125个,多态性比率平均为87.08%,不同引物扩增出的清晰条带数为5~11,平均为6.94条。多态性比率变幅为66.67%~100%,差异较大。
14份扁蓿豆的SSR扩增结果见图1和表3,18对SSR引物共扩增到136个条带,SSR扩增带的分子量为70~300bp,其中多态性条带109个,多态性比率平均为80.09%。不同引物扩增出的清晰条带数为3~13,平均每个引物扩增出的片段条带数为6.06条。不同引物扩增的多态性也存在较大的差异,每个引物扩增的多态性片段的百分率介于60.0%~100%。
Shannon指数和Nei’s指数既可以反映条带的丰富程度,又可以反映均匀程度,而丰富度与均匀度是衡量多样性的2个重要指标。Nei’s指数估算的18个ISSR引物和18对SSR引物扩增所得位点的种群内和种群间的基因多样性,不同位点对遗传多样性的贡献不同,SSR的Nei’s指数为0.284,平均Shannon指数0.438,种群内基因多样性为0.284,遗传分化系数为0.674,ISSR的Nei’s指数为0.352,平均Shannon指数为0.522,种群内基因多样性为0.352,遗传分化系数为1.000(表3)。
2.2 遗传相似性分析
SSR分析结果表明,利用18对SSR引物产生的109条DNA片段计算了供试材料间的遗传相似系数(GS)。SSR标记揭示的材料间GS值变化范围为0.669 7~0.885 3,SSR数据结果:遗传相似系数(GS)最小的是来自内蒙古呼和浩特市的6号黄花扁蓿豆材料和内蒙古通辽市3号黄花扁蓿豆材料(0.669 7),遗传距离最远,遗传相似程度最低;最大的是来自内蒙古包头市达茂旗的14号扁蓿豆材料和来自内蒙古赤峰市的12号扁蓿豆材料(0.885 3),遗传距离最近,遗传相似程度最高。
ISSR分析结果表明,利用18个ISSR引物产生的125条DNA片段计算了供试材料间的遗传相似系数(GS)。ISSR标记揭示的材料间GS值变化范围为0.447 6~0.811 2,ISSR数据结果:遗传相似系数(GS)最小的是来自内蒙古呼和浩特市武川县13号的扁蓿豆材料与内蒙古锡林郭勒盟锡林浩特市的9号扁蓿豆材料(0.447 6),遗传距离最远,遗传相似程度最低;最大的是来自内蒙古包头市达茂旗的14号扁蓿豆材料与内蒙古呼和浩特市的6号黄花扁蓿豆材料(0.811 2);遗传距离最近,遗传相似程度最高。2种标记所得结果基本相似。
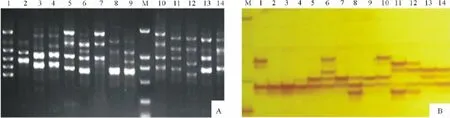
图1 UBC854(A)及 MTIC272(B)扩增产物电泳图谱Fig.1 Amplification results of UBC854(A)and MTIC272(B)
2.3 聚类分析
2.3.1 ISSR分析 根据以上对遗传相似性的计算结果,获得供试材料间的遗传距离GD(GD=1-GS),利用UPGMA法进行聚类分析,结果见图2,阈值在0.646将14份材料分为4大类。第一大类来自内蒙古呼和浩特市武川县13号的扁蓿豆材料;第二大类包括来自内蒙古赤峰市的12号扁蓿豆材料及来自内蒙古通辽市的11号扁蓿豆材料;第三大类包括来自内蒙古海拉尔市的2号细叶扁蓿豆材料、内蒙古包头市的4号黄花扁蓿豆材料、内蒙古呼和浩特市的8号黄花扁蓿豆材料、内蒙古呼和浩特市的6号黄花扁蓿豆材料、内蒙古包头市达茂旗的14号扁蓿豆材料、内蒙古通辽市3号黄花扁蓿豆材料、内蒙古通辽市5号黄花扁蓿豆材料、内蒙古呼和浩特市的7号黄花扁蓿豆材料;第四大类包括内蒙古锡林郭勒盟白音锡勒的1号细叶扁蓿豆、内蒙古锡林郭勒盟锡林浩特市的9号扁蓿豆材料、内蒙古锡林郭勒盟西苏旗的10号扁蓿豆材料。

表3 ISSR和SSR标记的结果比较Table 3 Comparison of usefulness between ISSR and SSR markers
2.3.2 SSR分析 阈值在0.7将14份材料分为4大类(图3)。第一大类来自内蒙古锡林郭勒盟白音锡勒的1号细叶扁蓿豆;第二大类包括来自内蒙古海拉尔市的2号细叶扁蓿豆材料、内蒙古呼和浩特市的6号黄花扁蓿豆材料、内蒙古呼和浩特市的8号黄花扁蓿豆材料、内蒙古通辽市5号黄花扁蓿豆材料、通辽市的11号扁蓿豆材料、内蒙古通辽市7号黄花扁蓿豆材料、内蒙古呼和浩特市武川县13号扁蓿豆材料、内蒙古锡林郭勒盟锡林浩特市的9号扁蓿豆材料、内蒙古赤峰市的12号扁蓿豆材料、内蒙古包头市达茂旗的14号扁蓿豆材料、内蒙古锡林郭勒盟西苏旗的10号扁蓿豆材料;第三大类是来自内蒙古通辽市的3号黄花扁蓿豆;第四大类是来自内蒙古包头市土右旗的黄花扁蓿豆。

图2 14份扁蓿豆材料的ISSR聚类图Fig.2 Dendrogram of 14 M.ruthenicagermplasm by ISSR
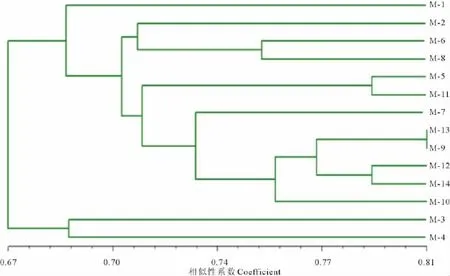
图3 14份扁蓿豆材料的SSR聚类图Fig.3 Dendrogram of 14 M.ruthenicagermplasm by SSR
2.4 2种标记的Mantel检验
通过Mantal测验进行表型性状欧式遗传距离矩阵、SSR和ISSR标记的遗传距离矩阵之间的相关性分析(图4)。结果表明,二者距离矩阵之间存在显著相关,相关系数r=0.019 6,t=0.121 2证明重要的表型性状能够较准确的反映出亲缘相近的材料的遗传变异。
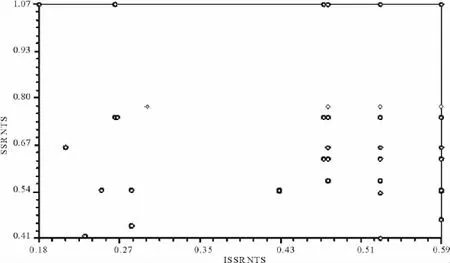
图4 扁蓿豆材料ISSR分析和SSR分析的关系Fig.4 The data relationship between ISSR and SSR of M.ruthenica
3 讨论与结果
分子标记的应用性评价是开展遗传多样性分析的基础工作之一[19]。由于所用标记技术和材料的不同,得出的结论不尽一致。扁蓿豆因其优异的特性具有很大的利用潜力。本研究对收集到的不同生态型的扁蓿豆资源进行了SSR和ISSR遗传多样性的分析,2种标记均表现出较高的多态性,表明2种标记在扁蓿豆植物种间分析是有效的。基于SSR和ISSR标记的遗传相似性系数及UPGMA聚类显示分析的综合结果来看,黄花扁蓿豆种质资源遗传基础较广,主要集中在同一个大的类群中,具有一定的同源性,亲缘关系较近;ISSR标记具有比SSR标记更高的PPB值,说明前者比后者具有更高的标记效率和有更多的多态性位点,说明SSR标记在亲缘物种间具有高度多态性,适于亲缘关系较近种群的分析[20]。因此认为ISSR标记在14份扁蓿豆材料间的扩增位点多态性主要由类群间的差异引起,SSR标记可高效的检测扁蓿豆种质资源中具有较近亲缘关系的种群的遗传多样性[21]。2种标记的结果相对一致,Mental测验二者距离矩阵相关系数为r=0.019 6,t=0.121 2。SSR 标记和ISSR标记的综合聚类分析表明,2种标记可以针对不同的类群和不同育种目标对野生扁蓿豆资源进行有目的的改良和利用,从而扩大扁蓿豆遗传基础,为扁蓿豆育种积累材料。本研究方法也可为今后扁蓿豆种质资源的创新利用和遗传多样性研究提供更为合理准确的参考。
扁蓿豆分布环境的多样性,导致扁蓿豆不同居群之间以及同一居群不同个体之间形态性状变异的多样性。扁蓿豆为苜蓿属中的一个野生种,是改良和育成新品种的巨大基因库,利用分子标记开展多基因聚合育种是未来育种的前途所在,借助截形苜蓿等模式牧草的分子标记研究成果,有助于尽快获得豆科牧草的大量标记和高密度图谱。
[1] 宁布,杜一民,陈山.内蒙古扁蓿豆属植物研究草地生物多样性保护研究[M].呼和浩特:内蒙古大学出版社,1995.
[2] 李鸿雁,李志勇,米福贵,等.利用微卫星标记鉴定扁蓿豆种质资源[J].华北农学报,2008,23(3):67-71.
[3] 李鸿雁,米福贵,宁红梅,等.扁蓿豆遗传多样性的SSR分析[J].中国草地学报,2008,30(2):34.
[4] 李志勇,师文贵,李鸿雁,等.内蒙古3种不同生态型扁蓿豆遗传多样性与亲缘关系的分析[J].西北植物学报,2009,29(12):2421-2428.
[5] 李鸿雁,李志勇,王小丽,等.内蒙古扁蓿豆遗传多样性的ISSR分析[J].西北植物学报,2011,31(1):52-56.
[6] 王照兰,杨持,赵丽丽,等.扁蓿豆不同品系ISSR标记遗传差异和遗传多样性[J].中国草地学报,2010,32(1):11-17.
[7] 田青松,韩冰,杨劼,等.96份雀麦属材料遗传多样性的ISSR分析[J].中国草地学报,2010,32(1):18-25.
[8] 刘美华,李忠超,陈海山,等.昆仑锦鸡儿的遗传多样性研究[J].广西植物,2005,25(1):53-57.
[9] 卢萍,赵萌莉,韩国栋,等.内蒙古小花棘豆遗传多样性的ISSR分析[J].西北植物学报,2007,27(6):1102-1107.
[10] 刘永财,孟林,张国芳,等.新麦草种质遗传多样性的ISSR分析[J].华北农学报,2009,24(5):107-112.
[11] Martin J P,Sanchez-Yelamo M D.Genentic relationships among species of the genus Diplotaxis(Brassiceae)using inter simple sequences repeat markers[J].Theoretical and Applied Genetics,2000,101:1234-1241.
[12] Mac Ritchie D,Sun G L.Evaluating the potential of barley and wheat microsatellite markers or genetic analysis of Elymus trachycaulus complex species[J].Theoretical and Applied Genetics,2004,108:720-724.
[13] 孙建萍,袁庆华.利用微卫星分子标记研究我国16披碱草遗传多样性[J].草业科学,2006,23(8):40-44.
[14] 谢文刚,张新全,马啸,等.中国西南区鸭茅种质遗传变异的SSR分析[J].草业学报,2009,18(4):138-146.
[15] 刘志鹏,杨青川,呼天明,等.用SSR标记研究不同耐盐特性四倍体紫花苜蓿的遗传多样性[J].作物学报,2006,32(4):630-632.
[16] 郭强,时永杰,魏臻武,等.河西走廊14种锦鸡儿遗传多样性SSR分析[J].草地学报,2007,16(3):229-233.
[17] Bernadette J,Sandrine F,Philippe B.Const ruction of two gengtic linkage maps in cultivated tet raploid alfalfa(Medicago sativa)using microsatellite and AFLP markers[J].BMC Plant Biology,2003,9(3):12-19.
[18] Mantel N.The detection of disease clustering and a generalized regression approach[J].Cancer Research,1967,27:209-220.
[18] 冯亮亮,唐红,李毅,等.甘肃红砂不同种群遗传多样性的ISSR分析[J].草业学报,2011,20(1):125-130.
[19] 王普昶,赵丽丽,易津,等.内蒙古野生华北驼绒藜种群遗传多样性和遗传分化研究[J].草业学报,2011,20(4):287-292.
[20] 乔岩,王汉宁,张成,等.玉米胚乳突变基因ae连锁累赘的SSR分析[J].草业学报,2011,20(1):140-147.
[21] 李鸿雁,李志勇,米福贵,等.中国扁蓿豆遗传多样性的表型和SSR标记分析[J].西北农林科技大学学报,2011,39(8):1-9.
