不同放牧制度对荒漠草原地上生物量及种间关系的影响
2012-08-20方楷宋乃平魏乐安慧
方楷,宋乃平*,魏乐,2,安慧
(1.宁夏大学西北土地退化与生态恢复国家重点实验室培育基地,宁夏 银川750021;2.宁夏大学学术期刊中心,宁夏 银川750021)
草原是我国面积最大的陆地生态系统,它的健康稳定对我国牧区的经济发展和全国生态安全具有重要意义。但中国90%的天然草原发生了不同程度的退化,其中严重退化草地近1.8亿hm2,严重威胁我国生态安全和牧区经济的发展。为此国家在多个牧区实施了围栏禁牧政策,以便实现退化草地系统自我恢复。大量研究表明,国家的围栏封育政策成效显著[1-5],但也有研究指出,长期封育不利于促进并提高草场的放牧利用价值,封育时间过长还会引起群落地上生物量逐渐降低、凋落物增加,抑制植物的再生和幼苗的形成,阻碍草原的繁殖更新[6-8]。长期围栏和过度放牧均不利于草地生态系统的健康[9,10],而合理调控放牧制度才能有利于草地生态系统功能稳定,实现利用和保护的有机衔接[11-15]。草地生产力是草地生态系统健康的核心指标,也是群落功能最重要的体现载体。以往的研究指出,适度放牧通过加速营养循环、改善冠层辐射状况、提高牧草光合能力、促进资源再分配等使牧草补偿或超补偿性生长[16];不同生育期的牧草对放牧的敏感性有所差异,合理调控放牧时期能够提高草场的放牧抗性[17];划区轮牧将草地划分为若干轮牧小区,按照一定次序逐区采食,适度的放牧间隔有利于牧草恢复生长[18]。生物量的变异系数(CV)反映了群落初级生产力的稳定性[19],研究放牧系统中生物量稳定性的变化情况对维持草地生态系统的可持续发展具有重要意义。草地植物群落中放牧造成的植被演替过程必定伴随着种间关系的变化,深入探究种间关系对不同强度、不同方式放牧干扰的响应能够为阐明放牧演替的原因和机制提供理论依据。
荒漠草原是荒漠与典型草原间的缓冲地带,由于生态环境的严酷性和气候的波动性,生态系统十分脆弱,具有发生荒漠化的潜在危险[20]。但是一直以来,对于生产力水平相对较高的典型草原和草甸草原研究较为系统,干旱荒漠草原植被因其种类贫乏、群落稳定性差,较少受到重视。因此,以围封9年的荒漠草原为研究对象,通过设置划区轮牧、自由放牧、围栏封育3种放牧制度以及轻、中、重3种放牧强度,对比分析不同放牧处理间牧草地上生物量的变化,明确该区域草畜平衡的临界点;通过在各放牧小区布设活动围笼,研究不同放牧制度对牧草地上生物量空间变异以及种间关系的影响,为荒漠草地合理利用提供理论依据。
1 材料与方法
1.1 研究区自然概况
研究区位于宁夏盐池县四墩子行政村的荒漠草原上,北纬 37°04′~38°10′,东经 106°30′~107°47′,该区域生态环境脆弱,常年风沙活动频繁,属于典型的中温带大陆性气候。年平均气温7.7℃,1月份平均气温-8.9℃,7月份平均气温22.5℃,≥0℃年积温为3 430.3℃,≥10℃年积温为2 949℃;年日照时数2 867.9h,日照率65%;年平均降水量250~350mm,大部分集中在6-8月,且多暴雨,降水年变率较大;年蒸发量2 136 mm,是降水量的6~8倍;年无霜期162d。大地貌为缓坡丘陵,地带性土壤为灰钙土,还有大面积的风沙土、黄绵土等非地带性土壤,土壤质地以沙壤、粉砂壤和沙土为主。主要植物种有:本氏针茅(Stipa bungeana)、赖草(Leymus secalinus)、白草(Pennisetum centrasiaticum)、牛枝子(Lespedeza potaninii)、蒙古冰草(Agropyron mongolicum)、糙隐子草(Cleistogenes squarrosa)、甘草(Glycyrrhiza uralensis)、丝叶山苦荬(Ixeris var.graminifolia)、刺叶柄棘豆(Oxytropis aciphylla)、老瓜头(Cynanchum komarovii)等。
研究区2010年5月1日—10月2日降水162.42mm,属于丰水年;7月平均气温26.35℃,高于往年平均气温(图1)。该数据来源于距离四墩子村约10km的自动气象站。
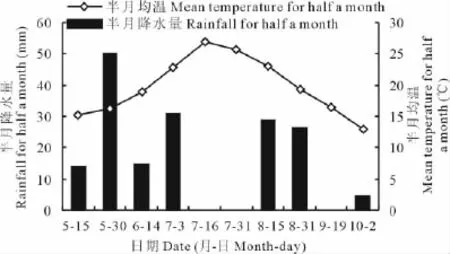
图1 半月降水量和半月均温Fig.1 Rainfall and mean temperature for half a month
1.2 试验设计
试验设置划区轮牧(RG)、自由放牧(CG)和封育禁牧(CK)3种放牧制度,其中划区轮牧、自由放牧的放牧强度分别为0.5只/hm2(轻)、0.75只/hm2(中)和1只/hm2(重)。轮牧区采用四区轮牧方式,每个轮牧小区放牧15d;自由放牧区实行连续放牧。选择地形、土壤和植被状况较为一致的草地88hm2(不含公共牧道),用围栏围成16个等面积的小区,每个小区5.5hm2。其中,轮牧小区12个、自由放牧小区3个、封育小区1个。针对荒漠草原自身较高的植被异质性,在每个放牧小区内随机放置5个活动围笼,围笼大小2m(长)×2m(宽)×1m(高),用以对比分析牧草地上生物量的时空变异及种间关系对不同放牧制度的响应。轮牧区在羊只转栏的同时将围笼移动到下一放牧小区;自由放牧区每30d移动1次围笼。选择健康无体病体重相近的2龄滩羊母羊,按上述放牧方式和强度分为6组进行放牧试验,组间羊只体重差异不显著(P>0.05)。放牧时间为2010年5月15日—2010年9月15日。
1.3 测定方法
放牧开始前(5月初),对各试验小区进行本底调查,结果显示:小区间生物量差异不显著(P>0.05)。放牧开始后,对各放牧处理按照不同周期进行植被调查(1m×1m的样方),轮牧区、自由放牧区、围封区的测定周期分别是15,30和15d,每次测定5个重复。植被样方调查按照活动围笼在放牧区内随机放置的位置进行,同时测定围笼内和围笼外各植物种的多度、高度、频度和样方总盖度,然后齐地面刈割样方内所有植被地上生物量,于实验室将牲畜可食的优良牧草和不食的有毒有害植物分装后用75℃烘干至恒重测其干重,所有地上生物量均为牲畜可食的优良牧草。
1.4 数据分析
采用Excel 2003进行数据的录入、整理和初步分析;采用SPSS 17.0进行各放牧处理间牧草地上生物量的单因素方差分析、多重比较和LSD检验(最小显著性差异,P<0.05);通过分别计算活动围笼内和围笼外牧草地上生物量的变异系数来对比分析不同放牧制度对荒漠草原草地生产力稳定性及其变异性的影响;采用国际通用的CANOCO 4.5软件分别对围笼内和围笼外植被样方中各植物种的密度进行PCA排序,探讨放牧对植物种间关系的影响。
2 结果与分析
2.1 放牧对牧草地上生物量的影响

表1 自由放牧区不同放牧强度牧草地上生物量的差异比较Table 1 Aboveground biomass in continuously grazed plots under different grazing intensities
2.1.2 划区轮牧制度下放牧强度对牧草地上生物量的影响 表2显示了划区轮牧制度下,不同放牧时期牧草地上生物量在3种牧压梯度下的方差分析和多重比较结果。其中,轻度牧压下,轮牧Ⅳ区的牧草地上生物量显著高于轮牧Ⅱ区、Ⅲ区;重度牧压下,轮牧Ⅱ区和Ⅳ区的牧草地上生物量显著高于轮牧Ⅲ区;而中度牧压下4个轮牧小区的牧草地上生物量差异不显著。比较相同放牧时期不同放牧强度的牧草产量发现:轮牧Ⅰ区和Ⅳ区在3种牧压下差异不显著,轮牧Ⅱ区和Ⅲ区差异显著。多重比较表明,轮牧Ⅱ区的牧草地上生物量在轻度和重度牧压之间差异显著,轮牧Ⅲ区则是中度牧压显著高于重度牧压。结合整个生长季的气象资料(图1),该地区2010年雨热不同期现象明显,即植被在生长季旺期遭遇了高温干旱胁迫环境,而生长季初期的雨水条件充足,这可能是引起轮牧Ⅱ区和Ⅲ区生物量在3种牧压梯度下产生差异的主要原因,并且放牧干扰后的环境胁迫作用使轮牧Ⅲ区的生物量比其他小区都低,同时也说明中度轮牧草场具有较高的抗胁迫能力。比较轮牧Ⅲ区和Ⅳ区发现,放牧干扰之后遭遇胁迫环境比在胁迫环境下放牧对牧草产量的影响更大,原因是干旱高温环境不仅作用于植被,还影响了羊只对草场的采食强度。
2.1.3 放牧制度对牧草地上生物量的影响 在同一放牧强度下,自由放牧区的年均牧草地上生物量均大于4区轮牧的(图2)。其中,在轻度牧压(F=40.418,Sig.=0.000)和中度牧压(F=11.284,Sig.=0.001)下,两者差异达到极显著水平。自由放牧方式下,牧草年均地上生物量随着放牧强度的增加而降低,轮牧区则相反,生物量随着放牧强度的增加而逐渐升高,并且自由放牧区牧草年均地上生物量在牧压梯度下的变化速率更大。自由放牧方式下,牧草年均地上生物量在3种放牧强度下差异显著(F=4.664,Sig.=0.016),轮牧区则未达到显著水平。对自由放牧区不同牧压梯度下牧草地上生物量进行多重比较发现,年均地上生物量在轻度牧压和重度牧压之间差异显著。以上结果表明,牧草地上生物量对不同放牧方式的响应不同,自由放牧较4区轮牧表现出更高的牧草生产力,而在牧压梯度下4区轮牧比自由放牧的草地生产力稳定性要好,从牧草地上生物量在牧压梯度下的变化趋势来看,4区轮牧能够比自由放牧耐受更大的放牧强度,这对缓解畜牧量增加与生态平衡之间的矛盾具有重要作用。

表2 轮牧区不同放牧强度牧草地上生物量的差异比较Table 2 Aboveground biomass in rotationally grazed plots under different grazing intensities
2.2 放牧对牧草地上生物量时空变异的影响
变异系数(CV)反映了年内牧草地上生物量的总体变异程度。一般认为,CV<0.1为弱变异,CV在0.1~1.0为中等变异,CV>1.0为强变异[21]。研究区年内牧草地上生物量在各处理下都属于中等变异程度,其中自由放牧区的变异系数在中度牧压下最小、重度牧压下最大,轮牧区的变异系数随着牧压梯度的增加而增加,并且2种放牧方式下的变异系数均比封育区小(表3),说明放牧干扰能够减小荒漠草原年内牧草地上生物量的时间变异。
生长季不同时期牧草初级生产力的变异系数对3种强度自由放牧响应不同(表4)。轻度和重度的自由放牧增加了生长季初期和末期牧草地上生物量的空间变异,降低了生长季旺期的空间变异,而中度自由放牧则刚好相反,它降低了生长季初期和末期牧草地上生物量的空间变异,增加其生长季旺期的空间变异。结合活动围笼内牧草地上生物量的变异系数可知,生长季初期、旺期、末期的牧草生产力稳定性分别在中度牧压、轻度牧压和中度牧压时最大。

图2 不同放牧制度牧草地上生物量在3种牧压梯度下的差异比较Fig.2 Mean aboveground biomass among different grazing systems

表3 不同放牧制度年内牧草地上生物量的变异系数Table 3 CV of the average biomass among different grazing systems
图3显示了4区轮牧方式下,活动围笼内外牧草地上生物量变异系数的年内动态曲线。围笼内牧草地上生物量变异系数在3种放牧强度下的动态变化规律基本相同,说明可以作为对照数据分析放牧对牧草生产力空间变异的影响,其波动性反映了不同植物种群在返青期、枯萎期等生物学特性上的差异以及干旱高温胁迫环境的影响。与活动围笼内相比,轻度轮牧对年内牧草地上生物量的空间变异降低明显,中度轮牧则略有增加,重度轮牧对其影响相对较小。
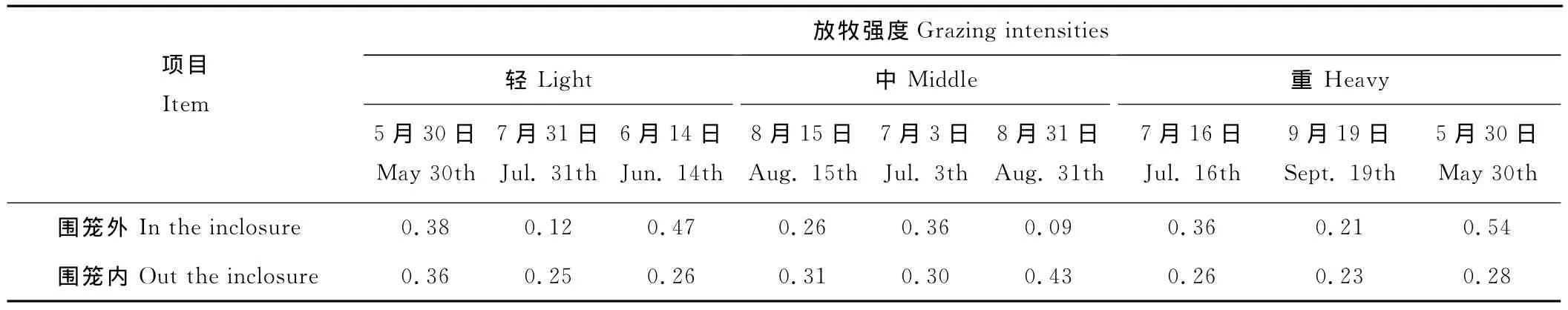
表4 自由放牧区围笼内外牧草地上生物量的变异系数Table 4 CV of aboveground biomass inner or outer the inclosure in continuously grazed plots
2.3 放牧对植被种间关系的影响
通过对样方中各植物种的密度进行PCA排序,对比分析不同放牧方式下活动围笼内外植被的种间关系,排序过程除去频度值小于20%的偶见种,排序图中各物种之间夹角的余弦值表明了种间关系特征。自由放牧区、轮牧区分别采用15个(笼内或笼外)和40个(笼内或笼外)样方进行排序,各排序图前两轴的特征值均大于75%,说明排序结果良好。通过植被调查发现,整个试验区呈现出3种植物群落(本氏针茅群落、赖草群落、白草群落)的斑块状镶嵌分布格局,从而认为本氏针茅、赖草、白草为研究区的建群植物。依据史志诚[22]主编的《中国草地重要有毒植物》一书,将刺叶柄棘豆、老瓜头和乳浆大戟归属于对畜牧业造成严重危害的毒草。根据退化草原恢复演替中植物种出现的时间及其在群落中的地位[23,24],认为糙隐子草是恢复演替中期优势植物,而铁杆蒿、牛枝子为恢复演替早期的优势植物。
在轻度自由放牧区,只有本氏针茅一种建群植物,原因是荒漠草原植被具有高度的异质性以及在样方调查过程中存在一定的随机性,并非放牧所致。对比围笼内外建群种与其他植物种之间的关系发现,放牧增加了本氏针茅与牛枝子、铁杆蒿的负相关性以及与刺叶柄棘豆的正相关性(图4)。同时,放牧还促进了一些新物种的生长,如乳白花黄芪、绵蓬和甘草等,说明轻度自由放牧能够降低建群种的优势度,为新物种的生长提供生存空间和所需资源。

图3 轮牧区围笼内外牧草地上生物量变异系数的年内动态曲线Fig.3 Dynamics of aboveground biomass’CV inner or outer the inclosure of rotationally grazed plots
在中度自由放牧区,围笼内外建群种之间的相关性变化很小,即赖草、白草、本氏针茅之间均表现为弱相关性或负相关性(图5)。放牧干扰也没有改变建群种与一些演替等级较低的植物种(糙隐子草、铁杆蒿、牛枝子、老瓜头、刺叶柄棘豆)之间的相互关系,说明中等强度的自由放牧不会引起种间关系的剧烈变化。
在重度自由放牧区,围笼内外3种建群植物之间的关系发生了较大变化,放牧增加了本氏针茅与白草的正相关性及与赖草的负相关性(图6)。与围笼内相比,围笼外糙隐子草与白草的负相关性减小,与赖草、本氏针茅的负相关性增大,牛枝子与本氏针茅的正相关性略有增加,铁杆蒿与本氏针茅和白草的负相关性增加,与赖草的负相关性减小。老瓜头与赖草在围笼内正相关,在围笼外负相关。放牧增加了刺叶柄棘豆与白草之间的负相关性以及与赖草之间的正相关性。重度自由放牧增加了本氏针茅群落与赖草群落之间的资源竞争,3种群落均表现出不同程度的逆向演替趋势,其中赖草群落受到的影响最大。
由于中英文在语言形式、语序结构、表达方式等方面存在较多差异,而合同中又常常使用长句来使得语义更加明确,因此在翻译长句时,常常出现逻辑混乱、语意不明的情况。这是因为中文结构在形式上较为松散,主要靠语义衔接,而英文更重视严密的逻辑关系和形式上的连接。在进行英译汉长句处理时,则经常打破英文行文的原有顺序,进行形式调整,以追求中文译文的准确、自然、流畅。在这些操作中,稍有不慎,就可能出现语句逻辑不清晰甚至混乱,造成译文让人感到费解的情况。因此,复杂长句的处理需要反复斟酌,慎之又慎,把每个成分之间的关系彻底理清再开始翻译。
在轻度轮牧区,围笼内外建群种之间的相关性变化较小,即围笼内外的赖草分别与白草和本氏针茅之间表现为弱相关性和强的负相关性(图7)。放牧干扰改变了糙隐子草与本氏针茅之间的相关性,两者在笼内负相关,笼外正相关;与围笼内相比,笼外牛枝子与赖草、白草的正相关性减弱,与本氏针茅的正相关性增强;铁杆蒿在围笼内与白草、本氏针茅正相关,与赖草负相关,在笼外刚好相反,其与白草、本氏针茅负相关,与赖草正相关;放牧增加了老瓜头和刺叶柄棘豆与赖草、白草的正相关关系。轻度牧压下的4区轮牧对研究区3种建群植物之间的关系影响较小,群落界限依旧明显,但是3种植物群落均表现出不同程度的逆向演替趋势。

图4 轻度自由放牧区围笼内外种间关系比较Fig.4 Interspecific relationships of inner or outer inclosure in continuously grazed plot with lax intensity
在中度轮牧区,围笼内外3种建群植物之间的相关性变化明显,放牧增加了本氏针茅与赖草、白草之间的正相关关系(图8)。糙隐子草在放牧作用下与本氏针茅的正相关性减弱,与白草、赖草的正相关性增加;围笼外牛枝子与本氏针茅、白草的负相关性较围笼内高;放牧干扰增加了铁杆蒿与赖草、白草的正相关性以及铁杆蒿与本氏针茅的负相关性;刺叶柄棘豆和老瓜头在笼内相伴随而出现,在笼外两者相关性较低;与笼内相比,放牧增加了老瓜头与白草和赖草的正相关性,减少了刺叶柄棘豆和本氏针茅群落的正相关性。中等强度的4区轮牧改变了研究区3种建群植物之间的关系,使群落间的界限逐渐模糊,缓解了群落界面上激烈的资源竞争关系,并且促进了本氏针茅群落的顺行演替,但是白草群落和赖草群落呈现出逆向演替的趋势。
在重度轮牧区,围笼内外3种建群植物之间的相关性基本相同,即赖草、白草、本氏针茅之间都表现为弱相关关系或者负相关关系(图9)。比较围笼内外糙隐子草与3种建群植物之间的关系发现,重度放牧增加了糙隐子草与本氏针茅的正相关性以及与白草和赖草的负相关性。牛枝子在围笼外与本氏针茅的正相关性也较围笼内高,老瓜头和刺叶柄棘豆与3种建群植物之间的关系变化较小。重度牧压下的4区轮牧对本氏针茅群落影响最大,长期干扰下将使其发生逆向演替,而白草群落和赖草群落的逆向演替趋势不明显。
综上所述,荒漠草原植物种间关系对不同放牧制度的响应各不相同。其中,中度自由放牧方式下围笼内外植物种间关系的差异最小,而重度自由放牧对植被的影响最大,不仅加剧了群落间的资源竞争,还会引起3种群落发生不同程度的逆向演替,中等强度的4区轮牧方式为研究区最优的放牧模式,它一方面减缓了群落界面上的资源竞争,另一方面促进了本氏针茅群落的顺行演替。

图5 中度自由放牧区围笼内外种间关系比较Fig.5 Interspecific relationships of inner or outer inclosure in continuously grazed plot with moderate intensity

图6 重度自由放牧区围笼内外种间关系比较Fig.6 Interspecific relationships of inner or outer inclosure in continuously grazed plots with heavy intensity

图7 轻度牧压轮牧区围笼内外种间关系比较Fig.7 Interspecific relationships of inner or outer inclosure in rotationally grazed plots with lax intensity

图8 中度牧压轮牧区围笼内外种间关系比较Fig.8 Interspecific relationships of inner or outer inclosure in rotationally grazed plots with moderate intensity
3 讨论
3.1 不同放牧制度下放牧强度对牧草地上生物量的影响
放牧系统中,牲畜采食、践踏等行为一方面通过伤害植物的正常组织、形态,抑制植物生长,另一方面通过移去植物顶端组织和衰老组织,刺激植物生长[25],而草地受损程度在其阈值范围以内可以通过自身的生理机制进行恢复,若超过该阈值则会发生退化。因此,放牧强度是抑制或促进草地系统持续健康稳定的最关键影响因子。与不放牧相比,合适的放牧强度能够促进植被的补偿或超补偿生长[15],从而提高草地系统的生产潜力。植物的补偿性生长与草地群落类型、放牧制度、放牧强度以及环境条件等密切相关[15,18,26-28]。结果显示,自由放牧制度下年均牧草地上生物量随着放牧强度的增加而逐渐降低,其中重度牧区较封育小区低;在光热水条件充足时(生长季初期),自由放牧小区的牧草地上生物量均比封育小区高,并且生物量随着放牧强度的增加而降低;当遭遇高温干旱胁迫环境时(生长季旺期),自由放牧小区牧草地上生物量的增长量均低于封育小区。

图9 重度牧压轮牧区围笼内外种间关系比较Fig.9 Interspecific relationships of inner or outer inclosure in rotationally grazed plots with heavy intensity
划区轮牧是我国天然草地放牧管理的发展方向[11],与传统的自由放牧制度相比,可提高植被的现存量、生长量和生产力[18],以及在干旱季节表现出较大的土壤呼吸速率[29]等。研究结果显示,在同一放牧强度下,自由放牧较4区轮牧草场的年均牧草地上生物量高,但是从地上生物量在牧压梯度下的变化趋势来看,4区轮牧能够比自由放牧耐受更大的放牧强度。划区轮牧通过人为控制使放牧期草地维持在可持续功能和健康所能承受的最低生理生长阈值以上,植被在再生长期逐渐恢复到放牧前的健康水平并产生一个生理生长的上限[11]。由于不同生育期的牧草对放牧的敏感性有所差异,牧草对放牧时期的响应也会不同。以往的研究指出,一般牧草在春季返青期和秋季结籽期较为脆弱,放牧抗性较低[11];晚秋放牧对来年春季菊苣(Cichoriumintybus)的生长有不利影响[30];不同植物种群的高度、盖度和密度对不同放牧时间的响应不一致,晚放牧能够有效提高主要植物种群的地上生物量[31]。本研究中,轮牧草场环境条件对牧草的影响较放牧时期更为明显,中等放牧强度的轮牧草场地上生物量稳定性最高,表现出较好的抗胁迫能力。
群落稳定性是草畜平衡最重要的体现,关于放牧的研究无一不涉及这个问题,比如:放牧系统中植物种群各属性的消长[32]、物种多样性的研究[33]、群落空间分布格局的变化[34,35]等。但是一直以来,小尺度上放牧对牧草净初级生产力时空变异的影响研究不多。结果表明,自由放牧和4区轮牧都可以减少荒漠草原年内牧草地上生物量的时间变异;但其空间变异较为复杂,其中自由放牧区,轻度和重度牧压增加了生长季初期和末期牧草地上生物量的空间变异,降低其生长季旺期的空间变异,中度牧压则与之相反;而在轮牧小区,轻度牧压对年内牧草地上生物量的空间变异降低明显,中度牧压则略有增加,重度牧压对其影响相对较小。
3.2 不同放牧制度下放牧强度对牧草种间关系的影响
草原植物存在避食性和耐牧性两种放牧抗性,避食性指降低被牧食的机率和强度等机制,耐牧性指植物被牧食以后刺激植物再生长的机制[24]。家畜的选择性采食能够改变牧草的竞争力,这是一个随放牧持续变化的动态过程,家畜因草地结构连续变化而不断变化其择食性[11]。一般来说,放牧强度小牲畜的选择性采食强,放牧强度大牲畜的选择性采食弱。因此,适度的放牧能够改善群落结构,提高系统稳定性。本研究中,建群植物本氏针茅、赖草、白草较其他草种生物量大且植株个体高,因此被牧食的机率也最大,其放牧抗性表现为耐牧性;糙隐子草的植株相对矮小,通过空间机制采取避食策略;铁杆蒿的自然高度较低、牛枝子具有匍匐的植物结构,加之两者通过构成机制降低其适口性,避食性较强。由此可见,当放牧干扰大于建群植物的耐牧性时,将剩余一部分光照、土壤水分、养分等资源,从而被动增加了一些矮生禾草的资源竞争力;随着放牧干扰强度的进一步增加,建群植物逐渐被矮生禾草所取代;长期高强度的放牧干扰又会降低以矮生禾草为建群种的群落竞争力,而后被一些适口性差、匍匐生长的植物所取代;以此循环,直到牲畜不可食的有毒有害物种成为建群植物为止。本研究结果显示,放牧不仅改变了植物种群之间的关系,还会改变群落间的关系,并且不同的植物群落对放牧方式及强度的响应各不相同。其中,中度轮牧能够缓解群落界面上的资源竞争,对本氏针茅群落的顺行演替具有促进作用,增加了赖草群落和白草群落的逆向演替趋势;重度轮牧却不利于本氏针茅群落的顺行演替;中度自由放牧区的种间关系及群落间的关系变化最小;重度自由放牧不仅加剧了群落间的资源竞争,还会引起3种群落发生不同程度的逆向演替。
[1] 郑翠玲,曹子龙,王贤,等.围栏封育在呼伦贝尔沙化草地植被恢复中的作用[J].中国水土保持科学,2005,3(3):78-81.
[2] 曹成有,邵建飞,蒋德明,等.围栏封育对重度退化草地土壤养分和生物活性的影响[J].东北大学学报(自然科学版),2011,32(3):427-451.
[3] 蒋德明,李明,押田敏雄,等.封育对科尔沁沙地小叶锦鸡儿群落植被特征及空间异质性的影响[J].生态学杂志,2009,28(11):2159-2164.
[4] 张凡,祁彪,温飞,等.不同利用程度高寒干旱草地碳储量的变化特征分析[J].草业学报,2011,20(4):11-18.
[5] 赵景学,祁彪,多吉顿珠,等.短期围栏封育对藏北3类退化高寒草地群落特征的影响[J].草业科学,2011,28(1):59-62.
[6] 左万庆,王玉辉,王风玉,等.围栏封育措施对退化羊草草原植物群落特征影响研究[J].草业学报,2009,18(3):12-19.
[7] Huang Z,Gutterman Y.Artemisia monosperma achene germination in sand:effects of sand depth,sand/water content,cyanobacterial sand crust andtemperature[J].Journal of Arid Environments,1998,38(1):27-43.
[8] Eldridge D J,Westoby M,Holbrook K M.Soil surface characteristics,microtopography and proximity to mature shrubs:effects on survival of several cohorts of Atriplex vesicariaseedlings[J].Journal of Ecology,1992,78(2):357-364.
[9] 赵哈林,大黑俊哉,李玉霖,等.人类放牧活动与气候变化对科尔沁沙质草地植物多样性的影响[J].草业学报,2008,17(5):1-8.
[10] Green D R.Rangeland restoration projects in western New South Wales[J].Australian Rangeland Journal,1989,11(2):110-116.
[11] 侯扶江,杨中艺.放牧对草地的作用[J].生态学报,2006,26(1):244-264.
[12] 邹雨坤,张静妮,杨殿林,等.不同利用方式下羊草草原土壤生态系统微生物群落结构的PLFA分析[J].草业学报,2011,20(4):27-33.
[13] 刘颖,王德利,王旭,等.放牧强度对羊草草地植被特征的影响[J].草业学报,2002,11(2):22-28.
[14] 卫智军,李霞,刘红梅,等.呼伦贝尔草甸草原群落特征对不同放牧制度的响应[J].中国草地学报,2011,33(1):65-70.
[15] 马红彬,谢应忠.不同放牧强度下荒漠草原植物的补偿性生长[J].中国农业科学,2008,41(11):3645-3650.
[16] Oesterheld M,McNaughton S J.Effects of stress and time for recovery on the amount of compensatory growth after grazing[J].Oecologia,1991,85:305-313.
[17] 李勤奋,韩国栋,敖特根,等.划区轮牧中不同放牧利用时间对草地植被的影响[J].生态学杂志,2004,23(2):7-10.
[18] 邢旗,双全,金玉,等.草甸草原不同放牧制度群落物质动态及植物补偿性生长研究[J].中国草地,2004,26(5):26-31.
[19] 杨殿林,韩国栋,胡跃高,等.放牧对贝加尔针矛草原群落植物多样性和生产力的影响[J].生态学杂志,2006,25(12):1470-1475.
[20] 李景平,刘桂香,马治华,等.荒漠草原景观格局分析——以苏尼特右旗荒漠草原为例[J].中国草地学报,2006,28(5):81-85.
[21] Yonker C M,Schimel D S,Paroussis E,et al.Patterns of organic carbon accumulation in a semiarid shortgrass steppe,Colorado[J].Soil Science Society of America Journal,1988,52:478-483.
[22] 史志诚.中国草地重要有毒植物[M].北京:中国农业出版社,1997.
[23] 李政海,裴浩,刘钟龄,等.羊草草原退化群落恢复演替的研究[J].内蒙古大学学报(自然科学版),1994,25(1):88-98.
[24] 汪诗平.草原植物的放牧抗性[J].应用生态学报,2004,15(3):517-522.
[25] Huhta A P,Hellstron K,Rautio P,et al.Grazing tolerance of Gentianella amarellaand other monocarpic herbs:why is tolerance highest at low damage levels?[J].Plant Ecology,2003,166(1):49-61.
[26] 汪诗平,王艳芬.不同放牧率下糙隐子草种群补偿性生长的研究[J].植物学报,2001,43(4):413-418.
[27] 张荣,杜国祯.放牧草地群落的冗余与补偿[J].草业学报,1998,7(4):13-19.
[28] 刘艳,卫智军,杨静,等.短花针茅草原不同放牧制度的植物补偿性生长[J].中国草地,2004,26(3):18-23.
[29] 徐海红,侯向阳,那日苏.不同放牧制度下短花针茅荒漠草原土壤呼吸动态研究[J].草业学报,2011,20(2):219-226.
[30] Li G D,Kemp P D,Hodgson J.Regrowth,morphology and persistence of Grassland Puna Chicory(Cichorium intybus L.)in response to grazing frequency and intensity[J].Grass and Forage Science,1997,52(1):33-41.
[31] 刘红梅,卫智军,闫瑞瑞,等.荒漠草原主要植物种群特征对不同轮牧时间的响应[J].内蒙古草业,2007,19(2):16-18.
[32] 闫瑞瑞,卫智军,杨静,等.短花针茅草原优势种群特征对不同放牧制度的响应[J].干旱区资源与环境,2008,22(7):188-191.
[33] 卫智军,杨静,杨尚明.荒漠草原不同放牧制度群落稳定性研究[J].水土保持学报,2003,17(6):121-124.
[34] 刘振国,李振清.不同放牧制度下冷蒿种群小尺度空间格局[J].生态学报,2004,24(2):227-234.
[35] 许清涛,黄宁,巴雷,等.不同放牧强度下草地植物格局特征的变化[J].中国草地学报,2007,29(2):7-12.
