黄河源区高寒草原的植被退化与土壤退化特征
2012-08-20周华坤赵新全温军陈哲姚步青杨元武徐维新段吉闯
周华坤,赵新全,温军,2,陈哲,2,姚步青,杨元武,徐维新,4,段吉闯,2
(1.中国科学院西北高原生物研究所,青海 西宁810008;2.中国科学院研究生院,北京100049;
3.青海大学农牧学院草业科学系,青海 西宁810016;4.青海省气象局,青海 西宁810001)
黄河源区是我国青藏高原重要的水源涵养区,黄河干流总径流量的49%源于该地区[1,2]。黄河源区也是青藏高原最主要的畜牧业基地之一,草地资源丰富,牧草品质优良,是发展草地畜牧业的物质基础[1]。黄河源区的草地类型主要包括高寒草甸、高寒草原、高寒灌丛及高寒沼泽等,分布面积占江河源区总土地面积的54.04%[2]。迄今为止,黄河源区退化草地约有357.13×104hm2,占草地总面积的1/3,其中严重退化草地面积约95.66×104hm2,占退化草地面积的26.79%[3],这已经威胁到当地的生态环境、生物多样性保护和畜牧业经济的发展[4]。为保护黄河源区的生态环境,恢复治理退化草地,需要充分了解草地退化演替过程中的一些生物学过程和特征。然而,黄河源区的草地研究多限于高寒草甸的结构、功能研究及其对退化的响应[3-6],而对高寒草原的研究报道相对较少[7],这不利于退化高寒草原的恢复治理和管理调控。同时,高寒草原作为黄河源区最重要的草地类型,其退化状态和程度直接决定着这一地区在涵养水源、调节气候等方面所具有的生态潜力。为此,以黄河源区高寒草地中最具代表性的紫花针茅(Stipapurpurea)高寒草原为对象,在青海省果洛州玛多县对高寒草原退化演替过程中的有关土壤特性、植物群落、地上地下生物量进行了详细研究,以期揭示高寒草原退化过程中的植被-土壤特点,草地生产力和生态系统功能之间的关系等,为退化高寒草原恢复治理提供科学依据并为其退化程度诊断提供一些量化指标。
1 材料与方法
1.1 野外调查与取样
根据研究区域草地退化程度,采用天然草地退化、沙化、盐渍化的分级指标[8]和北方草地退化四级梯度标准[9],同时参考已经发布的“高寒草原退化程度的界定方法”青海省地方标准[10],利用空间分布代替时间演替的方法[11]来研究植物群落演替动态和土壤特征的变化。本研究以此划分并选择不同退化程度的高寒草原样地(表1),共4处,依次为未退化高寒草原、轻度退化高寒草原、中度退化高寒草原和重度退化高寒草原,每类样地面积均为30m×40m,土壤为高山草原土。根据玛多县农牧局20世纪80年代的草地综合调查[12],研究样地原先植被均为典型的未退化紫花针茅高寒草原植被,土壤为高山草原土,此后由于过牧、鼠害及暖干化气候等原因,出现了演替进程中退化程度不等的草地类型。
2009年9月采用样方法[13]在上述四类样地内分别随机选择6个样方(100cm×100cm)进行植物群落调查,调查参数包括样方内植物群落和优势植物种的盖度、高度,然后按禾草、莎草和杂草三大经济类群分种齐地面采集植物地上部分,烘干后称量其生物量。另将枯草归为一类;在剪去植物地上部分后,采用土柱法[5]分4层(每层5cm)采集植物地下部分,细水将泥土冲洗干净后烘干根系并称量其生物量。

表1 取样地基本情况Table 1 The basic situation of plots
按照内蒙古草原土壤退化的进程和评价指标[14],同时测定了各个样地的土壤特征参数。在各样地内采集0~20cm表层土壤,6次重复,取混合样,经过预处理后分别进行pH值、铵态氮、硝态氮、速效磷、速效钾、全氮、全磷、全钾,有机质的测定[15],其中pH值用电位测定法;铵态氮用靛酚蓝比色法;硝态氮用紫外分光光度法,速效磷用硫酸钼锑抗比色法;速效钾用火焰光度法;全氮用重铬酸钾硝化蒸馏法;全磷用钼锑抗比色法;全钾用火焰光度法;有机质采用重铬酸钾氧化-稀释热法。另外,0~20cm表层土壤含水量的测定用时域反射TDR-200水分仪,0~30cm土壤紧实度用SC-900数显式土壤紧实度仪测定,每个样地内重复测定20次。
1.2 数据分析
生物多样性指数[16]的计算包括:
(1)S=n。“n”是样方中的物种数 (丰富度指数);

(3)Pielou指数(均匀度指数)。E1=H/lnS;
相似性指数[17]按照如下公式计算:

其中,R是样方x与样方y之间的相似性程度,xi和yi分别是植物种i在样方x和样方y中的相对地上生物量,X和Y分别是样方x和样方y中所有植物种相对地上生物量的总和。R的变化范围为0~1.00。
牧草质量按张大勇等[18]提出的草场质量指数(index of grassland quality,IGQ)来评价。牧草按其适口性划分为5类(优、良、中、差、毒),适口性值依次为3,2,1,0,-1。
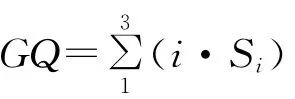
试验数据采用SPSS 17.0统计软件进行统计分析。
2 结果与分析
2.1 植被退化过程
2.1.1 植物群落地上生物量、物种多样性特征和草地质量 典型紫花针茅草原退化过程中,中度退化阶段的地上生物量显著大于轻度和未退化阶段的地上生物量,而后者显著大于重度退化阶段的地上生物量,但未退化和轻度退化阶段的地上生物量差异不显著(表2);紫花针茅地上生物量占总生物量的比例随着高寒草原退化而持续降低(表2)。草地质量指数和优良牧草地上生物量比例也随着草地退化加剧而降低(表2)。这与黄河源区退化高寒草甸[7]、藏北退化紫花针茅高寒草原[19]的规律是一致的。到重度退化阶段,可食优良牧草已经很难找到,未退化草地中的优势植物种针茅属植物已经消失,草地质量下降极其显著(P<0.01)。

表2 不同退化程度高寒草原植物群落特征和草地质量(n=6)Table 2 Properties and qualities of plant community at different degraded alpine steppe
植物群落物种丰富度、多样性指数和均匀度指数在紫花针茅高寒草原中度退化阶段最高,随着退化程度加大,呈单峰式曲线变化规律(表2),这与不同植物种群在退化演替过程中的消长变化密切相关。在中度退化阶段物种数最高,这与草地退化导致大量杂类草蔓延滋生有关;未退化高寒草原上,物种数目最少,优势种单一,这与生草层厚度等因素有关。原有的研究认为,高寒草原的退化与过度放牧关系密切[7,19],由于牧草在营养价值和采食难易程度等方面的差异,家畜有择食性,可显著改变植物种间竞争格局,引起物种侵入或迁出以及群落组成变化,这是本研究中不同退化程度下高寒草原生物多样性变化的直接原因。高寒草原植物种在退化演替系列中一般可以分为3类:减少种,如紫花针茅、溚草(Koeleriacristata);中立种,如二裂委陵菜(Potentillabifurca)、梭罗草(Roegneriathoroldiana)和苔草(Carextristachya)等;增加种,如矮火绒草(Leontopodiumnanum)、披针叶黄花(Thermopsislanceolata)和沙生风毛菊(Saussnreaarenaria),这也可以从植物群落优势种的更替变化中得到证实(表2)。所以,从生物多样性的角度出发,草原的保护应有食草动物的适当放牧,草原植物资源的科学经营应以高植物多样性和高生产力为目标。
2.1.2 植物群落的高度和盖度 高寒草原植物群落分上下两层,上层主要是紫花针茅、垂穗披碱草(Elymus nuntans)等禾本科植物,随草地退化加剧,植物群落的上层高度逐渐降低(表3);下层主要是一些杂类草植物,如矮火绒草、二裂委陵菜、狗娃花(Heteropappushispidus)、细叶亚菊(Ajaniatenuifolia)、沙生风毛菊等,整体而言高度都较低(小于5cm),在紫花针茅高寒草原的退化过程中,植物群落下层高度的变化不显著,无规律可循(表3),表明下层杂类草高度的变化与高寒草原退化程度关系不密切,这与典型高寒草原的特性有关[20]。
随着草地退化程度加大,禾草盖度和莎草盖度逐渐下降,以重度退化阶段的盖度最低。群落总盖度、杂草盖度呈先增加后减小的变化规律,其中以中度退化阶段最高,杂草盖度以未退化高寒草原最低(表3)。这表明在高寒草原逆行演替的过程中莎草与禾草在群落中处于竞争劣势,而杂草的大量入侵决定着退化阶段的草地群落特征。
2.1.3 植物群落间相似性指数 植物群落之间的相似性指数能够表示不同退化草地的植物组成及其地上生物量分配的差异[20]。如果不同草地植物组成及其地上生物量分配相似,则它们间的相似性指数就接近1。未退化高寒草原和轻度退化高寒草原之间的相似性指数最大,和重度退化高寒草原的相似性指数最小(表4)。随着高寒草原之间的退化程度差异增加,植物群落之间的相似性指数逐渐减小。
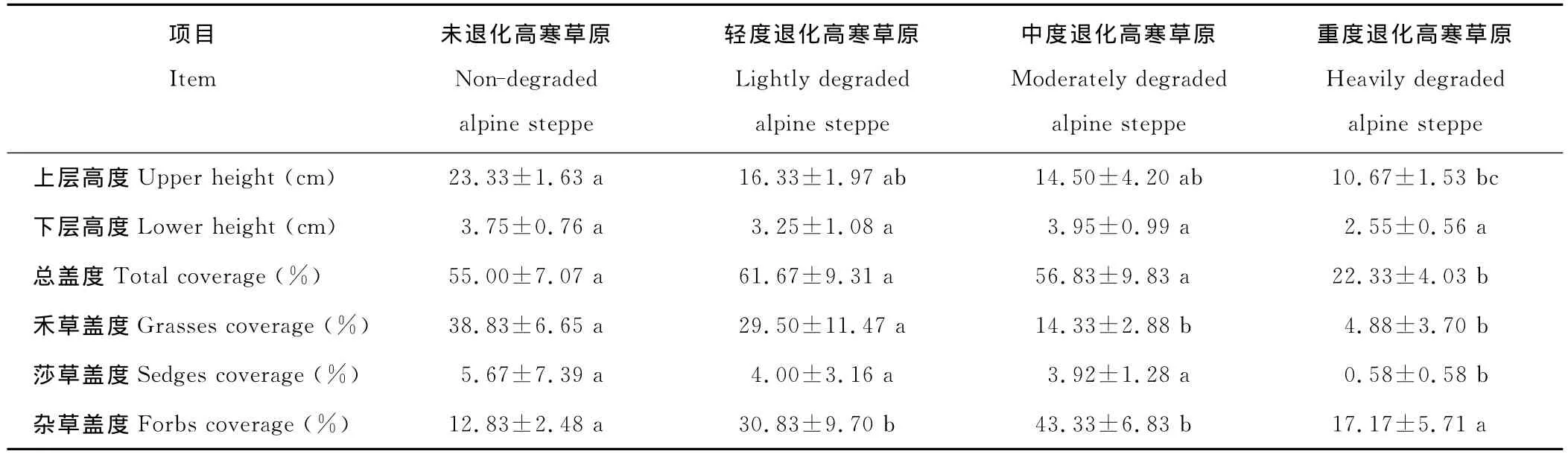
表3 不同退化程度高寒草原植物群落高度和盖度特征 (n=6)Table 3 The height and coverage of plant community at different degraded alpine steppe
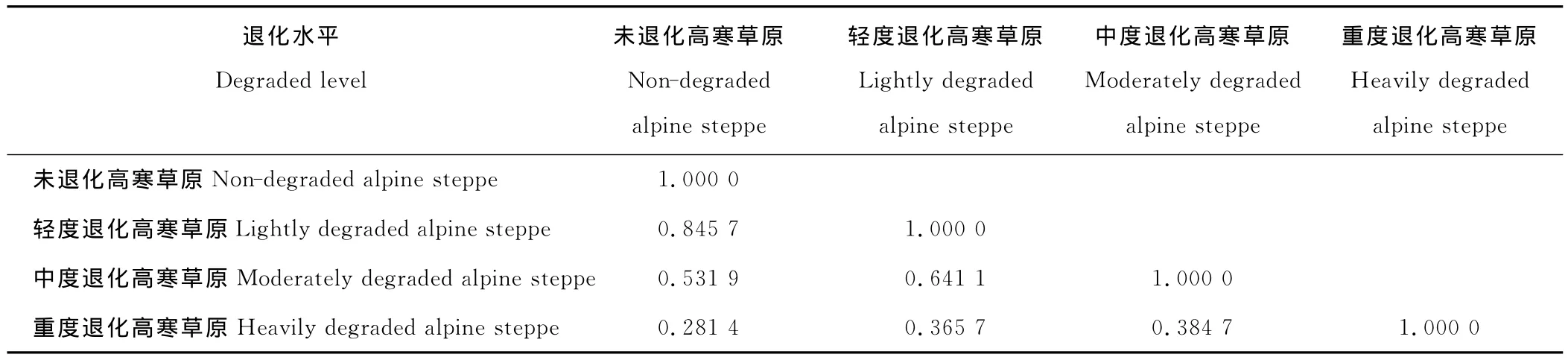
表4 不同退化程度高寒草原植物群落间相似性指数Table 4 The similarity indices of plant communities at different degraded alpine steppe
2.2 不同退化阶段地上地下生物量的变化
2.2.1 地上生物量的变化 中度退化阶段高寒草原地上总生物量最高,而在重度退化阶段最低(表2),不同经济类群植物地上生物量的消长变化有所不同(图1)。随着退化程度的加大,禾草绝对生物量一直下降,且彼此间差异显著(P<0.05);莎草地上生物量从未退化阶段到中度退化阶段变化不明显,重度退化阶段略有下降,但不显著(P>0.05);杂类草地上生物量的变化与莎草和禾草的变化趋势有所不同,呈倒“V”型变化趋势,以中度退化阶段为最高,显著高于其他各个阶段(P<0.01),其他3个阶段杂草生物量相似,差异不显著(P>0.05)。
莎草地上生物量比例的变化与其绝对量变化规律基本相同(图2),从未退化阶段到中度退化阶段变化不明显,重度退化阶段略有下降,但不显著(P>0.05)。禾草和杂类草地上生物量相对比例的变化趋势正好相反(图2),均与其绝对量的变化趋势不一致(图1)。未退化和轻度退化高寒草原的禾草生物量比例显著高于中度退化和重度退化高寒草原(图2,P<0.05),而未退化和轻度退化高寒草原的杂类草生物量比例显著低于中度退化和重度退化高寒草原(图2,P<0.05),莎草地上生物量比例与禾草和杂草的变化趋势截然不同,在不同退化阶段的草地群落中并未出现剧烈响应。因此,可以看出高寒草原退化演替的不同阶段,杂类草和禾草生长状况影响着整个群落结构和功能的变化,同时也决定了草地质量的高低。随草地退化的加剧,杂类草和禾草地上生物量比例呈一高一低的变化趋势,在由轻度退化向中度退化演替的某一时期,杂类草与禾草地上生物量比例相等(图2),这种此消彼长的现象说明在轻度退化和中度退化阶段高寒草原之间存在一个草地质量突变和衰退的阈值。
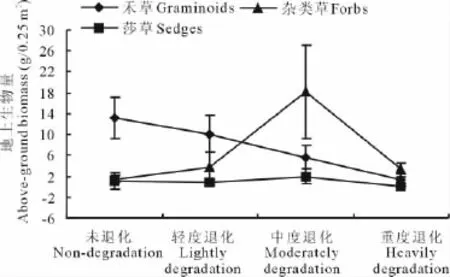
图1 不同退化程度高寒草原植物类群地上生物量Fig.1 Above-ground biomass of plant functional groups in different degraded alpine steppe
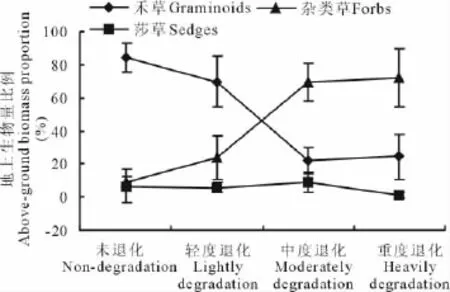
图2 不同退化程度高寒草原植物类群地上生物量比例Fig.2 Above-ground biomass ratio of plant groups in different degraded alpine steppe
2.2.2 地下生物量的变化 地下生物量的分层分布反映其垂直空间分布规律。随着高寒草原退化程度增加,地表0~20cm总生物量的变化趋势与地上生物量的变化规律一致,呈倒“V”型(表5),不过峰值出现在轻度退化的高寒草原,分别是未退化、中度退化和重度退化高寒草原的1.16,1.05和2.24倍。高寒草原地下生物量随着退化程度加剧,变化趋势与高寒草甸有所不同[5],这与不同草地类型的优势种生活型特征、生草层致密程度等有关系。各土层中植物地下生物量随着深度的增加而明显减小(表5),其中不同深度地下生物量之间差异显著;对于同一土层而言,不同退化程度高寒草原地下生物量之间不存在明显差异。不同退化程度高寒草原的地下生物量随土壤深度的分布关系都可以用对数函数y=a·lnx+b来描述,其相关性均达到极显著水平(表5,P<0.01)。
在不同退化程度的高寒草原中,0~20cm土层内地下总生物量的77%以上分布在0~10cm表层土壤中,0~15cm表土中的地下生物量占95%以上。可见植物根系的绝大部分分布在0~15cm土层中,这也是高寒草原地下部分分布模式的一个主要特点[1,2]。重度退化高寒草原分布在各层的植物根系量显著低于其他演替阶段的高寒草原,这反映了高寒草原退化演替与地下根系浅层化特点的相关性,这种反应特点与内蒙古退化草原有相同之处[21]。Coupland[22]综合国际生物计划期间12个生态定位站的资料,认为放牧干扰导致的草地退化具有使草地地下生物量向0~20cm集中的趋势,许多学者的研究表明高寒草原的退化与过重的放牧载畜量直接相关[7,19],本研究也证明了这一点。
2.2.3 地上地下生物量之间的关系 地上生物量与地下生物量的比率在中度退化阶段最大,未退化阶段次之,重度退化阶段最小,地上总生物量和地下总生物量之间也达到极显著正相关水平(P<0.01)(表6)。
2.3 物种多样性与生产力的关系
在植物群落尺度上,未退化、轻度退化和中度退化紫花针茅高寒草原的物种数目和地上生物量之间均呈“V”型变化规律,而重度退化高寒草原呈单峰变化规律,可用二次曲线较好的拟合(图3)。相关性分析表明,在未退化高寒草原地上生物量和物种丰富度之间的关系呈显著的正相关关系(r=0.743 9),轻度退化高寒草原地上生物量和物种丰富度之间的关系呈不显著的正相关关系(r=0.271 4),中度退化高寒草原地上生物量和物种丰富度之间的关系呈不显著的负相关关系(r=-0.593 7),重度退化高寒草原地上生物量和物种丰富度之间的关系呈显著的负相关关系(r=-0.711 9)。随着退化程度增加,高寒草原生产力与物种多样性的相关性发生了明显变化。
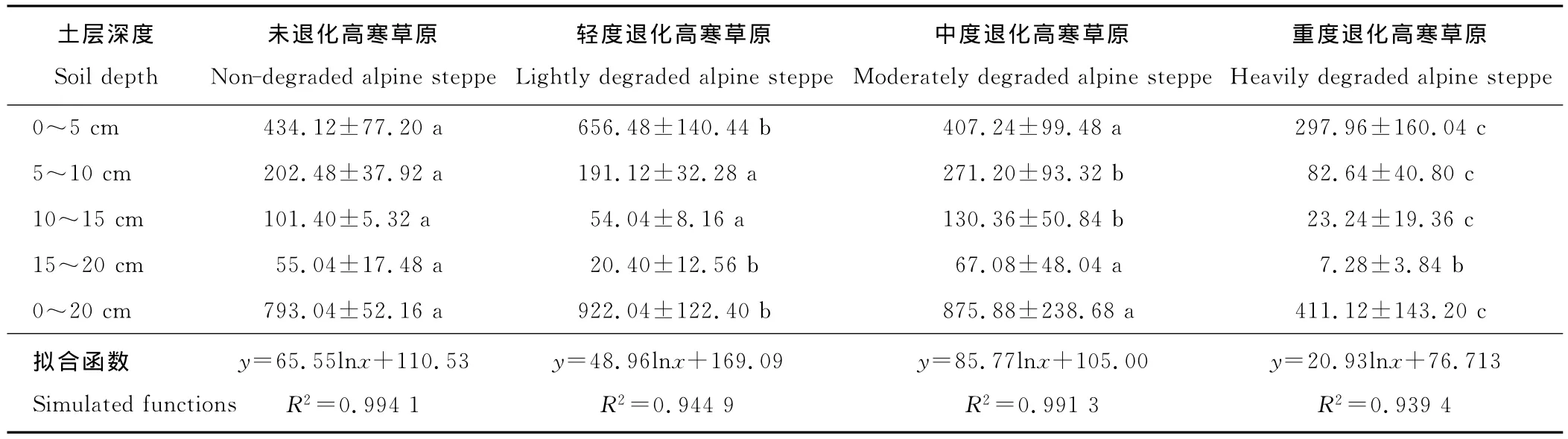
表5 不同退化程度高寒草原地下生物量分层分布Table 5 Layered distribution of underground biomass in different degraded alpine steppe g/m2

表6 不同退化程度高寒草原地上生物量与地下生物量及其关系Table 6 Above-ground biomass and below-ground biomass,and the relationship between them in different degraded alpine steppe

图3 不同退化阶段高寒草原物种丰富度与地上生产力的关系Fig.3 The relationship between the species richness with plant productivity in different degraded alpine steppe
生物多样性与群落生产力的关系属于群落生态学中的经典论题,揭示生产力对生物多样性的作用机制对于认识生物多样性的维持机制具有重要意义[23]。众多研究表明,物种多样性与生产力的关系表现为多种形式,如单峰型、线性正相关、线性负相关、U型以及不相关关系[23-26],影响多样性与生产力关系的因素很多,其中尺度被认为是重要的影响因素[25-30]。Gross等[26]利用长期生态系统监测网络的数据分析了不同生态尺度上物种多样性与生产力的关系,结果表明,大尺度上物种多样性与生产力之间呈单峰型关系;而在小尺度上(群落内)出现显著负相关关系。本研究中,重度退化、中度退化高寒草原生产力与物种多样性为负相关关系,与此结论是一致的。Schmid等[30]指出:当环境因素同生物多样性和生产力的关系都为正相关时,物种丰富度与生产力的关系为正相关关系。而当同一环境因素(如土壤肥力)对生物多样性和生产力的影响相反时,可能会导致物种丰富度与生产力的关系为负相关或峰型相关[31,32]。这与高寒草原退化演替过程中群落结构功能特征、土壤养分有效性和资源供给状态密切相关,以往的研究亦表明,高寒草原退化引起了土壤肥力等环境因子的变化[32],这使高寒草原物种丰富度与生产力的关系由显著正相关转变为负相关。

图4 高寒草原物种多样性指数与生产力的关系Fig.4 The relationship between the diversity index and productivity in alpine steppe
在区域尺度上,本研究中的高寒草原不论地上、地下生物量和总生物量,物种多样性与生物量之间都呈“V”型变化规律,可用二次曲线较好的拟合(图4),即较高和较低的生产力对应相对较高的物种多样性,而生产力居中时,对应的物种多样性指数较低,这与表2的数据规律是一致的。中国科学院海北高寒草甸生态系统研究站矮嵩草(Kobresia humilis)草甸、小嵩草(K.pygmaea)草甸和金露梅(Potentilla fruticosa)灌丛群落中物种多样性与生产力的关系呈线性增加关系,藏嵩草沼泽化草甸群落中线性增加关系不显著[33],与紫花针茅高寒草原物种多样性指数与生产力的关系不一致,这与高寒草地类型、草地受干扰状态、生物环境因子、尺度大小不同等有关系,在区域范围内,物种丰富度和植被生产力的关系主要是由环境变量驱动,如气候、土壤肥力和干扰程度[31,34-36]。相关性分析表明,在本研究区域紫花针茅高寒草原地上生物量和物种多样性之间的关系呈不显著的正相关关系(r=0.292 1),地下生物量与总生物量和物种多样性之间的关系均呈不显著的负相关关系(r=-0.072 95,r=-0.026 6)。
2.4 土壤退化
2.4.1 不同退化程度高寒草原土壤物理特征 土壤湿度不仅与蒸发蒸腾相互制约,而且还受降水、太阳辐射等因素的制约。随着紫花针茅高寒草原退化程度的加大,土壤湿度呈单峰式变化趋势,未退化和轻度退化高寒草原显著高于中度和重度退化高寒草原(表7)。高寒草原土壤湿度和植被盖度没有明显的对应关系,这和黄河源区退化高寒草甸的土壤含水量变化规律[5]有所不同,可能与区域降水量、太阳辐射、草地类型及土-草-根的耦合关系有关,有待进一步研究。
土壤紧实度受到土壤质地和结构性等的影响而变化。土壤愈疏松或是土壤中有大量的根孔、小动物穴或裂隙,则空隙度大而紧实度小;反之,土壤愈紧实[37]。随着高寒草原的退化程度加大,土壤紧实度逐渐减小(表7)。其中重度退化高寒草原土壤紧实度显著低于其他高寒草原,轻度和中度退化高寒草原显著低于未退化高寒草原,这与重度退化高寒草原植物稀疏,根系量锐减(表5),鼠类危害严重等密切相关。

表7 不同退化程度高寒草原土壤物理特征Table 7 Soil physical characteristics in different degraded alpine steppe
2.4.2 不同退化程度高寒草原土壤化学成分 土壤有机质、氮素和磷素等是土壤主要的养分指标,同时有机质还是形成土壤结构的重要因素,直接影响土壤肥力、持水能力、土壤抗侵蚀能力和土壤容重等,是土壤特性的重要指标之一[38],其变化状况可以指示土壤退化与否。
本研究中,随着高寒草原退化程度的增加,土壤表层中pH变化有总体上升趋势,且均高于8.0(表8)。土壤有机质在未退化和轻度退化阶段含量较高,中度退化和重度退化阶段较低,总体呈下降趋势(表8)。Dormaar等[39]的研究表明,放牧减少土壤有机质是草地重牧退化的重要表征。在本研究中,随着高寒草原退化到中度退化阶段,虽然植被覆盖度下降不显著(表3),但水土流失渐趋严重,有机质含量减少明显(表8),草土的耦合关系开始被扰动。表层土壤有机质含量受地上植被影响较大,退化草地表层枯草的数量增加,土壤中有机质含量在轻度退化条件下有所增加,这与蔡晓布等[32]对藏北紫花针茅高寒草原的研究结果是一致的。随着退化程度的加剧,生物量显著减少,所以有机质含量逐渐降低,而未退化草地土壤有机质含量稍低于轻度退化草地中,可能是由于未退化草地植物生长旺盛,对土壤有机质利用率较高的原因。

表8 不同退化程度高寒草原土壤化学成分Table 8 The soil chemical composition in different degraded alpine steppe
全氮量在各退化阶段变化不明显,相对而言,轻度和重度退化阶段较高,未退化和中度退化阶段较低(表8)。随着高寒草原退化程度的加剧,硝态氮含量减少趋势明显,其中重度退化高寒草原的硝态氮含量明显低于其他阶段,无法满足植物生长的需要,氨态氮的变化趋势与硝态氮正好相反(表8)。不同退化阶段高寒草原速效氮的变化与蔡晓布等[32]对藏北退化高寒草原的研究结果和黄河源区退化高寒草甸速效氮的变化[5]有所不同。
全磷含量在各退化高寒草原之间变化无明显规律可循,相对而言,在轻度退化阶段含量较高,其他退化阶段较低(表8)。随着高寒草原退化程度的加剧,速效磷含量减少趋势明显,其中重度退化高寒草原的速效磷含量明显低于其他阶段(表8),其变化规律与蔡晓布等[32]对藏北退化高寒草原的研究结果不同,速效磷含量明显低于高寒草甸[5],这可能与地域、取样和植被类型等有关系。速效磷与青海其他地区高寒草原土壤磷含量[40]相比,各退化阶段土壤磷素含量处于中等偏下水平,所以在退化高寒草原恢复治理过程中加施磷肥是有必要的。
土壤表层速效钾随着高寒草原退化程度加剧而减少,规律明显,其含量能满足植物生长对钾的需要,与藏北紫花针茅退化高寒草原的研究结果[32]一致。
植被退化是黄河源区高寒草原土壤退化的直接原因,而土壤退化也必然引起植被退化,二者互为因果。侯扶江等[41]认为草地退化不仅是植被和土壤的退化,也是2个子系统耦合关系的丧失和系统相悖所致。内蒙古典型草原在退化过程中首先表现为植被异质化,植被的异质化导致土壤某些元素的异质化[42]。由于土壤有较强的抗拒退化的能力,具有土壤稳定性的特点[43,44],所以在土壤退化之前植被的退化特征已经显现。高寒草原生态系统在适度放牧利用的情况下,能流和物流基本上处于平衡状态,生产水平比较稳定,土壤亚系统保持良好的结构,能够正常地发挥其功能,为植物生长提供所需要的空间、养分和水分,同时植物的残落物和家畜的排泄物归还给土壤,使草-畜-土维持良好的循环与平衡[1]。人为过度干扰是导致高寒草原出现逆向演替的主要因素[19,20]。最近几十年,紫花针茅高寒草原广泛分布的玛多县由于存在严重的超载过牧现象[7],牲畜过度啃食和践踏,草本植物的正常生长发育受到抑制,稳定的物质平衡受到破坏,草群变得低矮稀疏,组成结构改变,盖度减少,产草量下降,物种数和优良牧草量明显下降,与此同时,土壤也明显退化,有机质和速效氮含量下降,土壤紧实度同时下降,呈现退化和初始沙化的典型特征。本研究仅仅指出了有关高寒草原植被和土壤的退化特征,其退化驱动因素及其贡献率、退化机理、物种多样性和生产力关系及其调控机理仍然需要继续深入。
3 结论
随着高寒草原退化程度加大,植被盖度、草地质量指数和优良牧草地上生物量比例逐渐下降,草地间的相似性指数减小。而植物群落多样性指数和均匀度指数随着退化程度加大呈单峰式曲线变化规律,在中度退化阶段达到最高值。地上总生物量在中度退化阶段最高,在重度退化阶段最低。随着退化加剧,禾草地上生物量减少显著,杂草类植物地上生物量显著增加而后减少,而莎草科植物地上生物量的变化与草地退化程度的关系不明显;禾草地上生物量比例减少显著,杂草地上生物量比例增加显著,而莎草地上生物量比例在不同退化阶段变化不明显,且都比禾草与杂草的比值低。高寒草原植物地下根系表现出浅层化特点,分布在土壤表层0~20cm的植物根系随土壤深度的增加而减少,且随草地退化程度的加剧根系生物量变化呈倒“V”型。各类群地上地下生物量之间均为正相关,达到显著水平,地上生物量与地下生物量的比率在中度退化阶段最大,在重度退化阶段最小。高寒草原物种多样性与生物量之间呈“V”型变化规律,随着退化程度增加,物种丰富度与生产力的关系由显著正相关转变为负相关。
随着高寒草原由未退化和轻度退化阶段演替到重度和中度退化阶段,土壤含水量、土壤有机质、速效磷和硝态氮的含量都明显减小,硝态氮含量在重度退化阶段不能满足植物生长的需要。随高寒草原退化程度加大,有机质含量在表层土壤中流失严重。速效钾和土壤紧实度随着高寒草原的退化程度加剧而减小,氨态氮在重度退化阶段的含量高于其他阶段,而pH值均高于8,且变化不明显。
随着高寒草原植被的退化演替,土壤退化越来越严重,贫瘠化不断加剧,到重度退化阶段,旱生沙生植物出现,呈现沙化初始景观。
[1] 赵新全,马玉寿,周华坤,等.三江源区退化生态系统恢复及可持续管理化[M].北京:科学出版社,2011.
[2] 王根绪,李元寿,王一博,等.青藏高原河源区地表过程与环境变化[M].北京:科学出版社,2010.
[3] 贺有龙,周华坤,赵新全,等.青藏高原高寒草地的退化及其恢复[J].草业与畜牧,2008,(11):1-9.
[4] 赵新全,曹广民,李英年,等.高寒草甸生态系统与全球变化[M].北京:科学出版社,2009.
[5] 周华坤,赵新全,周立,等.青藏高原高寒草甸的植被退化与土壤退化特征研究[J].草业学报,2005,14(3):31-40.
[6] 韩立辉,尚占环,任国华,等.青藏高原“黑土滩”退化草地植物和土壤对秃斑面积变化的响应[J].草业学报,2011,20(1):1-6.
[7] 周华坤,周立,刘伟,等.青海省玛多县草地退化原因及畜牧业可持续发展[J].中国草地,2003,25(6):63-67.
[8] 苏大学,张自和,陈佐忠,等.天然草地退化、沙化、盐渍化的分级指标GB/T19377-2003[S].2004-1-8.
[9] 李博.中国北方草地退化及其防治对策[J].中国农业科学,1997,30(6):1-9.
[10] 周华坤,赵新全,马玉寿,等.高寒草原退化程度的界定方法DB63/T981-2011[S].青海省质量技术监督局发布,2011.
[11] Barbour M,Burk G,Pitts J H.Terrestrial Plant Ecology[M].London:The Benjamin Publishing Company,1980:222-233.
[12] Zhou H K,Zhao X Q,Tang Y H,et al.Alpine grassland degradation and its control in the source regions of Yangtze and Yellow Rivers,China[J].Grassland Science,2005,51:191-203.
[13] Walker M.Community baseline measurements for ITEX studies[A].In:Molau U,Molgard P.ITEX Manual,2nd ed[C].Copenhagen:Danish Polar Center,1996:39-41.
[14] 李绍良,贾树海,陈有君,等.内蒙古草原土壤退化进程及其评价指标的研究[J].土壤通报,1997,28(6):241-243.
[15] 中国土壤学会.土壤农业化学分析方法[M].北京:中国农业科技出版社,1999.
[16] 马克平,黄建辉,于顺利.北京东灵山地区植物群落多样性的研究II丰富度、均匀度和物种多样性指数[J].生态学报,1995,15:268-277.
[17] Horn H S.Measurement of“overlap”in comparative oecologica studies[J].American Naturalist,1966,100:419-424.
[18] 张大勇,王刚,杜国祯.甘南山地草原人工草场的演替[J].植物生态学与地植物学学报,1990,14(2):103-109.
[19] 蔡晓布,张永青,邵伟.藏北高寒草原草地退化及其驱动力分析[J].土壤,2007,39(6):855-858.
[20] 赵恒军.高寒草原紫花针茅草地植物群落基本特征研究[J].黑龙江畜牧兽医,2010,6:95-97.
[21] 王炜,梁存柱,刘钟龄,等.羊草+大针茅草原群落退化演替机理的研究[J].植物生态学报,2000,24(4):468-472.
[22] Coupland R T.Grassland Ecosystems of the World Ecological Studies[M].Springer-verlag Press,1979.
[23] Mittelbach G,Steiner C F,Scheiner S M,et al.What is observed relationship between species richness and productivity?[J].Ecology,2001,82:2381-2396.
[24] Waide R B,Willig M R,Steiner C F,et al.The relationship between primary productivity and species richness[J].Annual Review of Ecology and Systematics,1999,30:257-300.
[25] Guo Q F,Berry W L.Species richness and biomass:dissection of the hump-shaped relationships[J].Ecology,1998,79:2555-2559.
[26] Gross K L,Willig M R,Gough L,et al.Patterns of species density and productivity at different spatial scales in herbaceous plant communities[J].Oikos,2000,89:417-427.
[27] Chase J M,Leibold M A.Spatial scale dictates the productivity-biodiversity relationship[J].Nature,2002,416:427-430.
[28] Chalcraft D R,Williams J W,Smith M D,et al.Scale dependence in the species-richness-productivity relationship:the role of species turnover[J].Ecology,2004,85:2701-2708.
[29] 陈生云,刘文杰,叶柏生,等.疏勒河上游地区植被物种多样性和生物量及其与环境因子的关系[J].草业学报,2011,20(3):70-83.
[30] Schmid B,Hector A,Huston M,et al.The Design and Analysis of Biodiversity Experiments[M].Oxford:Oxford University Press,2002.
[31] Kahmen A,Perner J,Buchmann N.Diversity dependent productivity in semi-natural grasslands following climate perturbations[J].Functional Ecology,2005,19:594-601.
[32] 蔡晓布,张永青,邵伟.不同退化程度高寒草原土壤肥力变化特征[J].生态学报,2008,28(3):1034-1044.
[33] 王长庭,龙瑞军,王启基,等.高寒草甸不同草地群落物种多样性与生产力关系研究[J].生态学杂志,2005,24(5):483-487.
[34] Baer S G,Blair J M,Collins S L,et al.Soil resources regulate productivity and diversity in newly established tall grass prairie[J].Ecology,2003,84:724-735.
[35] Bai Y F,Wu J G,Pan Q M,et al.Positive linear relationship between productivity and diversity:evidence from the Eurasian steppe[J].Journal of Applied Ecology,2007,44:1023-1034.
[36] Kreft H,Jetz W.Global patterns and determinants of vascular plant diversity[J].Proceedings of the National Academy of Sciences USA,2007,104:5925-5930.
[37] 朱祖祥.土壤学[M].北京:农业出版社,1993.
[38] 鲁如坤.土壤-植物营养学[M].北京:化学工业出版社,1998.
[39] Dormaar J F,Smoliak S,Willms W D.Distribution of nitrogen fractions in grazed and ungrazed fescue grassland Ahorizons[J].Journal of Range Management,1990:43(1):6-9.
[40] 青海省农业资源区划办公室.青海土壤[M].北京:中国农业出版社,1997:280-285.
[41] 侯扶江,南志标,肖金玉,等.重牧退化草地的植被、土壤及其耦合特征[J].应用生态学报,2002,13(8):915-922.
[42] 程晓莉,安树青,李远,等.鄂尔多斯草地退化过程中个体分布格局与土壤元素异质性[J].植物生态学报,2003,27(4):503-509.
[43] 李绍良,陈有君,关世英,等.土壤退化和草地退化关系的研究[J].干旱区资源与环境,2002,16(1):92-95.
[44] 杨惠敏,王冬梅.草-环境系统植物碳氮磷生态化学计量学及其对环境因子的响应研究进展[J].草业学报,2011,20(2):244-252.
