Cd2 + 对小麦渗透性物质含量及籽粒产量的影响
2012-07-10李友军朱志勇
李友军,朱志勇,黄 明
(河南科技大学农学院,河南洛阳471003)
0 前言
Cd2+是植物生命活动的非必需元素,但却是毒性最强的重金属之一,比其他重金属具有更强的从土壤向植物迁移的能力,更易在植物体内富集,通过食物链进入动物或人体,可引起对人体骨骼、肾、肝、免疫系统和生殖系统的毒害作用及其致癌、致畸、致突变作用,严重威胁着人类健康及生命安全[1-3]。
随着环境污染的日趋严重,Cd2+污染已成为国内外学者关注的焦点[4-7]。近年来,关于Cd2+对水稻[8]、小麦[9]、玉米[10]、棉花[11]、白菜[12]等作物生长的危害及生理生化的影响都有不少的报道,但是这些报道一般多集中在作物的苗期,关于Cd2+对作物全生育期生长发育的影响报道较少。所以,研究镉对小麦灌浆过程中一些生理特性及最终产量的影响具有重要的科学意义。
小麦是世界上最主要的农作物之一,其栽培面积、总产量居各种作物的首位。在中国,小麦是仅次于水稻的第二大作物,其产量和品质是中国人民饮食水平的提高和饮食安全的保障的关键。目前,中国受重金属污染的耕地面积近2 000 万hm2,约占耕地总面积的1/5。因此,重金属污染已经成为中国小麦生产限制因子之一,在重金属污染现状还不能改善的前提下,提高小麦抗性成为保证小麦稳产、高产、安全的重要途径,其中,渗透调节作为表征植物抗性强弱重要生理机制越来越受到关注[11-14]。
渗透调节是植物适应干旱、盐渍、低温、重金属等胁迫的重要机制之一[15]。渗透调节物质主要分两大类:以K+为主的无机物和以脯氨酸、可溶性糖为主的有机物。脯氨酸是一种小分子的渗透物质,是一种溶解度高、水合度高的氨基酸,主要功能是提高原生质渗透压,防止和减少原生质水分散失,是最为有效的渗透调节物质之一。许多植物在受到逆境胁迫时都会积累大量的脯氨酸[16-17],脯氨酸可以作为一个抗逆指标。文献[18]通过对水稻研究显示,随着镉浓度升高,叶片的脯氨酸含量呈上升趋势;另有学者认为不同植物在胁迫下脯氨酸积累各不相同,有些植物的脯氨酸含量并不增加。文献[19]研究发现,大麦在NaCl 胁迫下脯氨酸含量下降。对于重金属胁迫对可溶性糖含量影响的报道也不尽一致,文献[20]认为,作物中可溶性糖含量随着重金属浓度升高而降低,而文献[21]却认为,植物体内的可溶性糖含量随着重金属浓度升高呈“N”字型的降-升-降的变化。从他们的研究结果可以看出:高浓度的重金属处理会使植物体内可溶性糖含量下降。本试验采用不同耐Cd2+性的两个小麦品种,设置3 个Cd2+处理水平,通过大田栽培的方式,研究Cd2+胁迫对小麦灌浆期游离脯氨酸、可溶性总糖含量及籽粒产量的影响,分析其与小麦耐Cd2+性的关系,以期为中轻度重金属污染区的小麦稳产、高产和安全生产提供一定的科学依据和理论基础。
1 试验材料与方法
1.1 试验材料
试验在河南科技大学开元校区进行,选取前期通过水培试验筛选的不同耐Cd2+性的两个小麦品种:新麦21(耐Cd2+性强)和百农矮抗58(耐Cd2+性弱)[22],Cd2+处理设置对照、低Cd2+处理和高Cd2+处理3 个水平,处理浓度分别为0 mg/kg(CK)、10 mg/kg(CL)、100 mg/kg(CH),供试镉试剂为分析纯CdCl2·2.5H2O,施于地表,然后翻土,使CdCl2·2.5H2O 均匀分布于0 ~40 cm 的耕作层之中。2009 年10 月21 日播种,随机区组排列,重复3 次,小区为3 m×3 m,基本苗2.4 ×106株/hm2。常规管理,氮肥50%(质量分数)作底肥,50%在拔节期随水追入,磷钾肥作底肥一次性施入。
1.2 试验田土壤理化性质
取试验田0 ~40 cm 的土壤测定土壤理化性质,土壤Cd2+本底值为0.18 mg/kg,土壤含水量为10.38%。其他理化性质见表1。
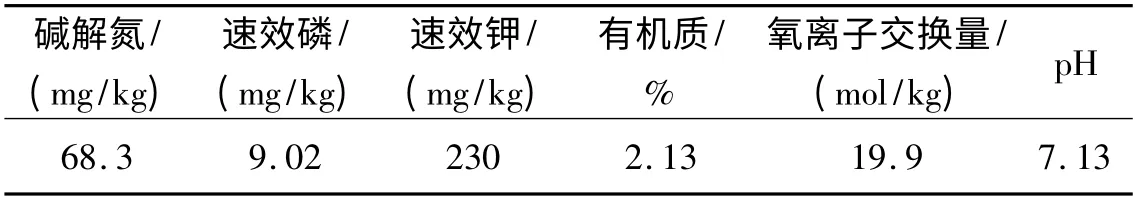
表1 试验田土壤的理化性质
1.3 取样方法
在各小区选择同天开花、表现一致且生长良好的穗挂牌标记足量,以后每隔5 d 取样1 次至收获,每区每次取20 个穗,105 ℃杀青后70 ℃下烘干,用于测定籽粒可溶性糖含量。取穗同时每小区取旗叶20 片,10 片烘干称重,用于测定可溶性总糖含量,另外10 片经液氮速冻30 min 后置于-70 ℃冰箱保存,用于游离脯氨酸含量的测定。小麦成熟后到收获前这段时间选取代表性植株10 株进行考种。
1.4 测定方法
旗叶和籽粒的可溶性总糖含量测定采用蒽酮比色法,游离脯氨酸含量测定采用酸性茚三酮比色法,产量去除边行和两头各50 cm 实收计产。
1.5 分析方法
采用DPS7.05 和Excel 分析试验数据。
2 结果与分析
2.1 Cd2+胁迫对两个小麦品种旗叶游离脯氨酸含量的影响
图1 为Cd2+胁迫对新麦21 和百农矮抗58游离脯氨酸含量的影响,由图1 可看出:两个小麦品种脯氨酸含量在整个灌浆时期呈上升趋势,其中,在开花期和花后5 d 时,新麦21 和百农矮抗58 的脯氨酸含量的顺序是CH >CL >CK;从花后5 d 开始,两个品种CH 的脯氨酸含量呈下降趋势,在花后10 d 以后新麦21 和百农矮抗58 的脯氨酸含量的顺序变为CL >CK >CH,且这个顺序一直保持到小麦成熟,这个顺序的改变说明脯氨酸含量未必与胁迫程度成正相关,严重胁迫条件下,脯氨酸含量可能降低。其原因可能是高Cd2+处理下,大量重金属积累,严重破坏了作物的细胞质膜,抑制脯氨酸的积累,使小麦自身的调节能力大大下降或者丧失。

图1 Cd2+胁迫对新麦21(左)和百农矮抗58(右)游离脯氨酸含量的影响
表2 为花后旗叶脯氨酸的平均净积累量,从表2 可以看出:两个小麦品种花后5 ~10 d 的脯氨酸净积累量都出现了负数,这可能与花后0 ~5 d 这段时间温度较高有关,高温胁迫一定程度上刺激脯氨酸的含量剧增;数据显示,CL 水平下新麦21 和百农矮抗58 的脯氨酸净累积量分别比各自CK 值提高了42.7%和20.6%,而CH 水平下却分别降低了49.4%和52.7%。由此可以得出两点结论:一是在轻度Cd2+胁迫下,脯氨酸的调节机制对小麦正常开花、灌浆起着重要的作用;二是脯氨酸累积量的高低作为反应作物Cd2+胁迫性强弱的重要指标。在此,试验验证了新麦21 耐Cd2+性大于百农矮抗58。
2.2 Cd2+胁迫对两个小麦品种旗叶可溶性总糖含量的影响
小麦灌浆过程中,叶片中贮存的光合产物开始大量向穗部运送,籽粒中的糖也大量转化为淀粉[23],旗叶是小麦重要的光合器官,旗叶中可溶性糖含量是籽粒可溶性糖含量、淀粉积累量及千粒重的主要影响因素之一[24],而旗叶可溶性总糖净积累量则反映了可溶性糖内在的合成速率与转化、运输速率。
图2 是镉胁迫下,两个小麦品种在灌浆过程中旗叶可溶性总糖含量变化趋势。两个品种在3 个处理下旗叶的可溶性糖含量均呈单峰曲线的变化趋势,旗叶可溶性总糖含量的高峰均出现在花后20 d。新麦21 旗叶中可溶性总糖含量在花后0 ~10 d 顺序为CL >CK >CH,而花后10 d 到成熟期顺序则为CK >CL >CH;百农矮抗58 旗叶可溶性总糖含量一直保持着CL >CK >CH。和CK 相比,CH 对旗叶可溶性总糖积累一直表现为抑制作用,高Cd2+处理使旗叶可溶性糖含量降低的原因可能是因为高Cd2+处理损害叶片膜系统,抑制了PSⅡ活性,导致其光合速率降低,使旗叶的光合同化产物积累量减少,或无足够的同化物转化形成其他蔗糖、可溶性糖等糖类,说明Cd2+胁迫破坏了小麦碳水化合物代谢途径[25-26]。
由图2 可知:整个灌浆过程中,在CK、CL、CH 这3 个水平下,新麦21 和百农矮抗58 旗叶中可溶性总糖净积累量的平均值分别为0.53 mg/d、0.31 mg/d、0.21 mg/d 和0.55 mg/d、0.62 mg/d、0.28 mg/d。方差分析显示,同一品种不同处理间的可溶性总糖净积累量差异性极显著(P <0.01),说明Cd2+胁迫抑制了小麦旗叶中可溶性糖的合成;在CL 或CH 水平下,不同品种的可溶性糖净积累量差异性极显著(P <0.01),百农矮抗58 净积累量大于新麦21 的净积累量,这可能是因为耐Cd2+性强的新麦21 旗叶中的可溶性糖往穗部的转化、运输速率比百农矮抗58 高。
2.3 Cd2+胁迫对两个小麦品种籽粒可溶性总糖含量的影响
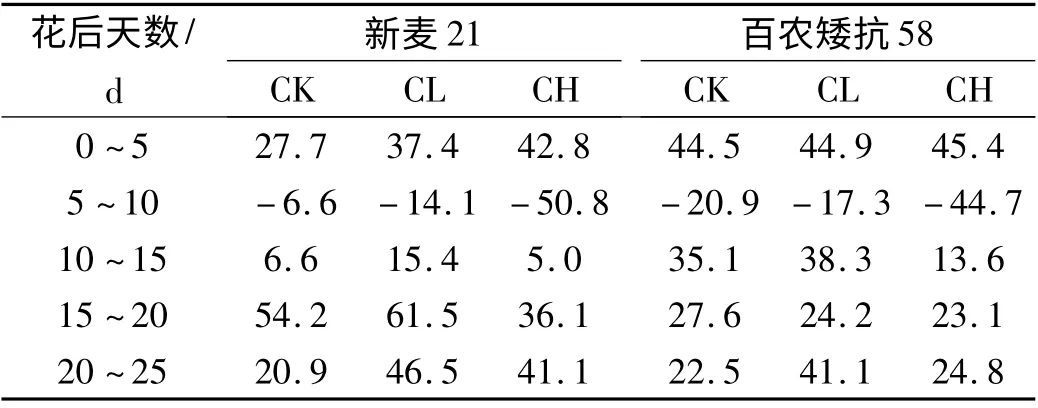
表2 花后旗叶脯氨酸的平均净积累量 μg/d
小麦籽粒中可溶性糖含量,一方面标志着叶源端的同化物供应能力;另一方面也能反映出籽粒对同化物的转化、利用能力[26]。图3 为镉胁迫对新麦21 和百农矮抗58 籽粒可溶性总糖含量的影响,从图3 可以看出:两个小麦品种籽粒中可溶性总糖含量在花后10 ~15 d 迅速下降,这是因为花后10 ~15 d 籽粒中大量的可溶性糖类转化为淀粉;花后10 d 到成熟期这段时间总体呈下降趋势,但是较平缓,这是因为花后20 d 以后,植株的光合能力开始下降,光合同化物含量下降,叶源端同化物供应能力逐渐降低,进而导致籽粒中可溶性糖含量下降。
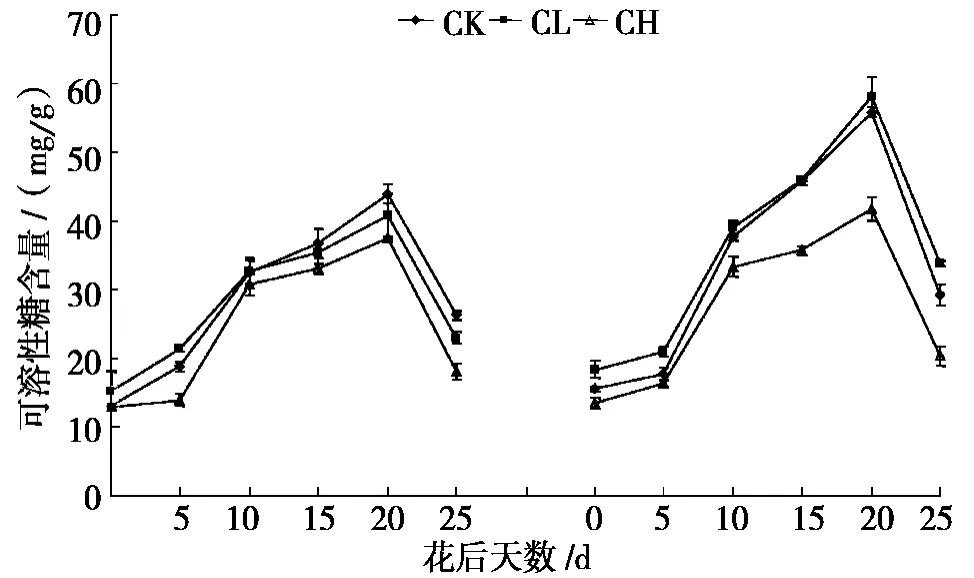
图2 镉胁迫对新麦21(左)和百农矮抗58(右)旗叶可溶性总糖含量的影响
从图3 中还可以看出:与CK 相比,CH 处理的籽粒可溶性糖积累受抑制,这可能是镉胁迫导致旗叶的可溶性糖往穗部运转受阻造成。数据显示:整个灌浆过程中,新麦21 和百农矮抗58 籽粒中的可溶性总糖含量平均值的相对值(相对值=镉处理值/对照值,相对值可以消除不同品种间固有的生物差异性)在CL 和CH 水平下分别为1.012、0.934 和0.919、0.910,相对值越大表示籽粒中可溶性糖积累量越高,所以镉胁迫下新麦21 籽粒中可溶性总糖积累量比百农矮抗58 高,这也恰好验证了镉胁迫下耐镉性强的新麦21 叶源端可溶性总糖转运速率比耐镉性弱的百农矮抗58 高这一观点。
经统计表明:新麦21 和百农矮抗58 各个处理在灌浆进程中籽粒可溶性糖平均日净积累量分别为12.2 mg/(g·d)、11.6 mg/(g·d)、11.1 mg/(g·d)和12.3 mg/(g·d)、9.3 mg/(g·d)、9.5 mg/(g·d),由于灌浆过程中籽粒中的大量可溶性糖转化为淀粉,所以本试验认为:镉胁迫下,高耐镉性新麦21 籽粒中可溶性糖转化为淀粉能力比低耐镉性的百农矮抗58 强。
2.4 Cd2+胁迫对两个小麦品种产量的影响
表3 为镉胁迫对两个小麦品种产量构成因素及产量的影响,从表3 可以看出:Cd2+胁迫通过影响小麦穗数、穗粒数和千粒重成产三因素进而影响小麦的产量。和CK 相比,CL 处理的有效穗数升高,CK 水平有效穗数则降低,这可能是因为低Cd2+处理促进小麦苗期的分蘖,而高Cd2+处理则使小麦幼苗分蘖受阻;CL 处理下新麦21穗粒数较CK 高,但是不显著,百农矮抗58 穗粒数显著降低,CH 处理下两个品种的穗粒数较CK都减少了;Cd2+处理对两个小麦品种籽粒千粒重都呈现为抑制作用,这可能是因为Cd2+胁迫抑制了籽粒中的糖类转化为淀粉的转化效率造成的,对百农矮抗58 抑制更显著,这也验证了Cd2+胁迫下高耐镉性新麦21 籽粒中可溶性糖转化为淀粉能力比低耐镉性的百农矮抗58 强的观点;分析最终产量可得出:新麦21 在CL 和CH 处理下的平均产量比CK 下降94.67 kg/hm2,百农矮抗58 下降了134.67 kg/hm2,Cd2+胁迫下耐镉性较强的新麦21 产量比耐镉性较弱的百农矮抗58 产量高。所以,在实际生产中,重金属轻度污染区可以通过选用耐性品种和农艺措施提高小麦穗粒数和千粒重,从而保证小麦高产稳产。
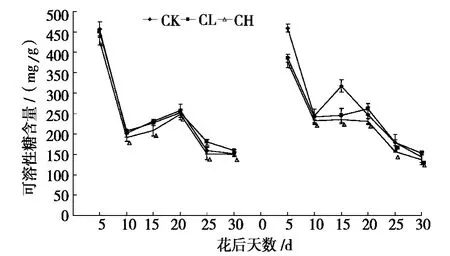
图3 镉胁迫对新麦21(左)和百农矮抗58(右)籽粒可溶性总糖含量的影响
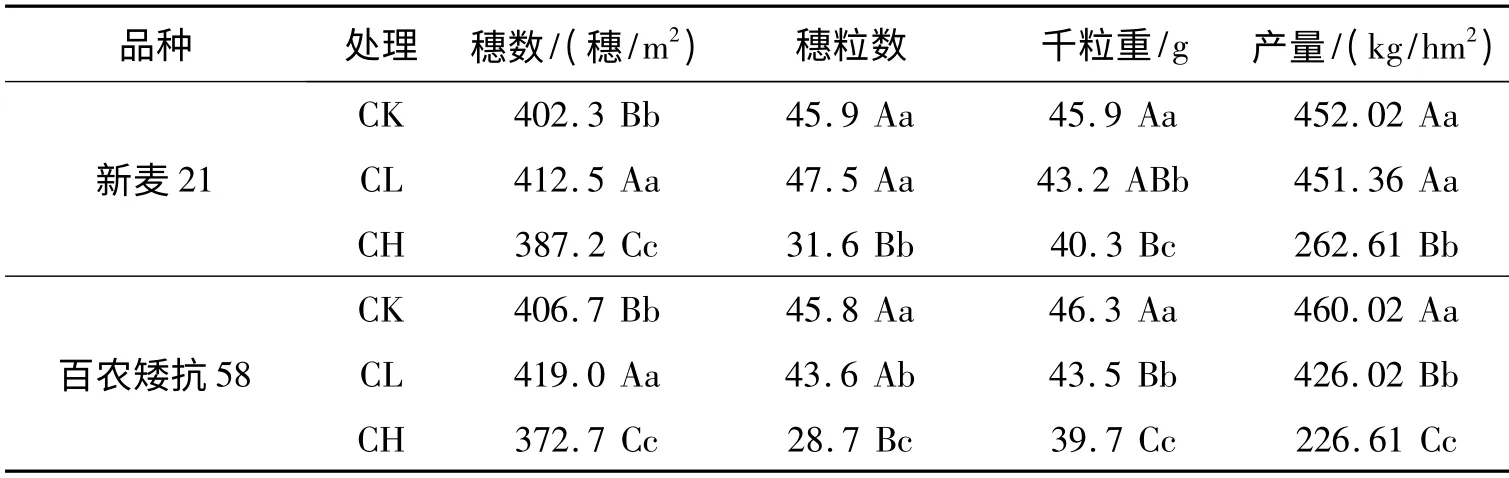
表3 镉胁迫对两个小麦品种产量构成因素及产量的影响
3 结论
(1)镉胁迫下,低镉浓度促进了叶片脯氨酸含量的积累,且随着灌浆过程的进行叶片的脯氨酸含量呈增加趋势,脯氨酸的调节作用保证了小麦轻度镉胁迫下能正常开花和灌浆;高镉浓度下由于大量重金属积累,严重破坏了小麦的细胞质膜,从而抑制脯氨酸的累积,影响了籽粒的正常灌浆;苗期耐镉性强的新麦21 在镉胁迫下表现出更强的自身调节作用。
(2)镉胁迫下,Cd2+破坏了小麦碳水化合物代谢途径,抑制了旗叶中可溶性糖的合成,阻碍了旗叶光合同化物往穗部的转运,进而影响了小麦籽粒可溶性糖的积累,同时镉胁迫抑制籽粒中可溶性糖转化成为淀粉;苗期耐镉性强的新麦21 在碳水化合物合成转运过程中,受破坏程度较轻。
(3)镉胁迫下,高镉浓度导致小麦减产,其中,苗期耐镉性强的新麦21 受影响程度较轻。
[1] Waisberg M.Molecular and Cellular Mechanisms of Cadmium Carcinogenesis[J].Toxicology,2003,192:95-117.
[2] Tudoreanu L,Phillips C J C.Modeling Cadmium Uptake and Accumulation in Plants[J].Advances in Agronomy,2004,84:121-157.
[3] 崔玉静,黄益宗,朱永官.镉对人类健康的危害及其影响因子的研究进展[J].卫生研究,2006,35(5):656-659.
[4] 郑喜珅,鲁安怀,高翔,等.土壤中重金属污染现状与防治方法[J].土壤与环境,2002,11(1):79-84.
[5] 朱志勇,郝玉芬,李友军,等. 镉对小麦旗叶叶绿素含量及籽粒产量的影响[J]. 核农学报,2011,25(5):1010-1016.
[6] Romic M,Romic D.Heavy Metals Distribution in Agricultural Topsoil in Urban Area[J].Environ Pollution,2003,43:795-805.
[7] 王凯荣.我国农田镉污染现状及其治理利用对策[J].农业环境保护,1997,16(6):274-278.
[8] 邵国胜,Muhammad J H,章秀福,等.镉胁迫对不同水稻基因型植株生长和抗氧化酶系统的影响[J].中国水稻科学,2004,18(3):239-244.
[9] 朱志勇,郝玉芬,吴金芝,等.镉胁迫对不同小麦品种以及Cd2+、Zn2+、Mn2+吸收和积累的影响[J].水土保持学报,2011,25(2):209-213.
[10] 孔祥生,郭秀璞,张妙霞.镉胁迫对玉米幼苗生长及生理生化的影响[J].华中农业大学学报,1999,18(2):111-113.
[11] 郑世英,张秀玲,王丽燕,等. 铅和镉胁迫对棉花种子萌发及有机渗透调节物质的影响[J]. 中国棉花,2007,34(5):16-26.
[12] 江帆,黄海泉,尹凤英,等.不同浓度镉对白菜幼苗生长受害与蛋白质组变化关系分析[J]. 农业环境科学学报,2008,27(6):2345-2350.
[13] 郭晓音,严重玲,叶彬彬.镉锌复合胁迫对秋茄幼苗渗透调节物质的影响[J].应用与环境生物学报,2009,15(3):308-312.
[14] 胡春霞,汤洁.重金属对蒲公英种子萌发及叶片渗透调节物质含量的影响[J].北方园艺,2008(11):27-30.
[15] Spickett C M,Smirnoff N,Ratdiffe R G.Metabolic Responses of Maize Roots to Hyperosmotic Ahock[J].Plant Physiol,1992,99:856-863.
[16] 朱虹,祖元刚,王文杰,等.逆境胁迫条件下脯氨酸对植物生长的影响[J].东北林业大学学报,2009,37(4):86-89.
[17] 汤章城.逆境条件下植物脯氨酸的累积及其可能的意义[J].植物生理学通讯,1984(1):15-21.
[18] 王阳阳,任艳芳,周国强,等.镉胁迫对不同抗性水稻品种幼苗生长和生理特性的影响[J].中国农学通报,2009,25(24):450-454.
[19] 杜志钊,邹磊,张艳敏,等.NaCl 处理对大麦种子萌发后生长量和相关生理生化指标的影响[J].上海农业学报,2010,26(2):34-37.
[20] 张义贤,张丽萍.重金属对大麦幼苗膜脂过氧化及脯氨酸和可溶性糖含量的影响[J].农业环境科学学报,2006,25(4):857-860.
[21] 丁晓辉,任丽萍,张春荣,等.Cd2+胁迫对紫花苜蓿叶绿素和可溶性糖含量的影响[J]. 华北农学报,2007,22(增刊):64-66.
[22] Li Y J,Zhu Z Y.A Study on Cd2+Resistance of Different Wheat Cultivars[C]//CMBB2010,IEEE.2010:210-216.
[23] 苏振刚,杨卫兵,田纪春,等.不同类型高产小麦植株、籽粒可溶性糖含量与其干物质积累的关系[J].中国农学通报,2007,23(9):307-311.
[24] 李友军,熊瑛,陈明灿,等.氮、磷、钾对豫麦50 旗叶蔗糖和籽粒淀粉积累的影响[J].应用生态学报,2006,17(7):1196-1200.
[25] 王贵民,陈国祥,陈利,等.镉胁迫下杂交稻2 品种幼苗的抗性及镉含量的差异[J].农村生态环境,2004,20(4):44-47.
[26] 薛香,梁云娟,韩占江,等.播期对百农矮抗58 小麦籽粒可溶性糖、淀粉积累的影响[J].河南农业科学,2010(7):15-17.
