部分牡丹品种的染色体核型分析
2012-04-07侯小改郭大龙张亚冰李雪萍张赞平
侯小改,郭大龙,张亚冰,李雪萍,张赞平
(河南科技大学a.农学院;b.林学院,河南洛阳471003)
0 前言
芍药属(Paeonia L.)牡丹组(sect.Moutan DC.)植物为中国特有,栽培品种也起源于中国[1-2],遗传资源十分丰富。中国牡丹在长期的栽培训化过程中,在株型、花型、花色等性状上产生了丰富多采的变异类型。但由于对其遗传背景、亲缘关系缺乏深入系统的研究,育种工作受到很大限制。
核型分析是分类学的一项重要依据[3],是探讨植物亲缘关系和系统演化的一种有效方法[4],有助于揭示物种遗传进化的过程和机制[5]。目前,牡丹野生种的核型分析多数已有报道,它们均为二倍体,属2A型核型,但某些种不同居群的核型有变化[6-15]。关于牡丹栽培品种的核型报道较少[16-20],但出现了不同产地品种的核型存在一定差异,甚至有的品种不同研究者得出的结果也不同的现象。形成这种差异的原因,可能与研究者的制片技术及分析方法有关,也可能与栽培条件或地理位置的变化有关。因此,本文对14个不同株高、花型及花色的牡丹品种核型进行了比较研究,以期为进一步探讨牡丹育种途径、杂交育种亲本选配及不同株型、花型、花色遗传规律等提供细胞学依据。
1 材料与方法
1.1 供试材料
试材来源于中国洛阳国家牡丹基因库,品种名称及特征见表1。
1.2 方法
2012年3月下旬采集0.5~1.0 cm饱满花蕾,用对二氯苯饱和溶液处理3~4 h后,蒸馏水冲洗干净,用卡诺氏固定液(φ(无水乙醇)∶φ(冰醋酸)=3∶1)固定24 h以上,固定好的材料如不能及时解离及制片,可将材料换至体积分数为70%酒精中,4℃冰箱保存备用。
将保存在冰箱中的材料用蒸馏水清洗,然后用1 mol/L的HCl在60℃解离5~10 min,蒸馏水漂洗,用改良的苯酚品红染色,压片法制片、镜检。
在显微镜下选择染色体分散较好的细胞进行染色体计数并照相。染色体计数和核型分析依据文献[21]的分析标准进行,每种材料均观察30个左右细胞进行染色体计数。分别选用5个分散良好的中期分裂相进行分析测量,得到核型数据。
染色体类型分析按照文献[22]的分类系统,核型类型分析参照文献[23]的分类标准。
染色体相对长度=染色体绝对长度/染色体组总长度×100%[21]。

表1 供试材料
染色体臂比值=染色体长臂长度/染色体短臂长度。
2 结果与分析
2.1 染色体核型的变化
所有供试材料的染色体数目均为2n=2x=10。各品种核型结构的主要参数见表2,各品种中期染色体及核型见图1。染色体着丝点类型分为:m代表中部着丝粒区;sm代表近中部着丝粒区;st代表近端部着丝粒区。
供试的14个品种可划分为3类核型。‘银鳞碧珠’、‘藕丝魁’、‘罂粟红’、‘紫红争艳’归为第1类,其核型组成完全相同,为2n=2x=10=6m+2sm+2st;‘酒醉杨妃’、‘红梅傲霜’、‘春归华屋’、‘金玉交章’、‘银粉金鳞’、‘十八号’、‘白玉’、‘肉芙蓉’归为第2类,其核型组成为2n=2x=10=8m+2st;‘万花盛’和‘佛门袈裟’归为第3类,其核型组成为2n=2x=10=8m+2sm。3类核型的对称性的由大到小的顺序为第3类,第2类,第1类。比较上述3类核型,其共同点是第1组、第2组、第3组均由中部着丝点染色体组成,所不同的是,第1类核型比第2类、第3类核型多了一对近中部着丝点染色体,而第3类与第1类、第2类染色体核型在第5组染色体上不同,不是近端部着丝点,而是近中部着丝点染色体。
依据文献[23]核型分类标准,供试品种均为2A类型,均属于对称型核型。根据文献[23-24]的观点,在系统演化上处于比较古老或原始的植物,往往具有较对称的核型,而不对称的核型则往往出现在较进化或特化的植物中。本文观察的14个品种中,从核型类型上看,‘银鳞碧珠’、‘藕丝魁’、‘罂粟红’、‘紫红争艳’可能属于较进化的类型。但是,这4个品种从株高、花色、花型等特征上无相近之处,第2类和3类亦如此。
2.2 染色体长度的变化
牡丹品种核型特征及核型图见表2和图1,表2和图1表明:‘紫红争艳’染色体最长,‘十八号’最短。上述牡丹品种染色体由大到小顺序为:‘紫红争艳’,‘肉芙蓉’,‘藕丝魁’,‘佛门袈裟’,‘银鳞碧珠’,‘银粉金鳞’,‘罂粟红’,‘金玉交章’,‘白玉’,‘酒醉杨妃’,‘万花盛’,‘红梅傲霜’,‘春归华屋’,‘十八号’。但是,牡丹各品种间染色体的大小及每个染色体组内各个染色体的大小,彼此相差较小,表现为连续性大小变化,均为大型染色体。且染色体大小与株高、花色及花型等性状间无明显相关性。
2.3 “随体”的变化
在‘十八号’、‘银粉金鳞’的第5对染色体短臂上及‘红梅傲霜’的第4对染色体短臂上有随体染色体存在。但这些随体染色体是不稳定的,即有的细胞中可见,有的则无。而在其他供试材料中未发现具有随体染色体。推测产生此种不稳定的原因可能是由于第Ⅳ组或第V组具近端部着丝点染色体短臂上带有较小的随体,两者往往相连而不易识别,只有在随体较延伸的细胞中方可辨认[17]。
3 讨论
3.1 关于核型类型
文献[16]观察了7个牡丹二倍体品种的核型,除‘青龙卧墨池’外均为2n=2x=10=8m+2st;文献[17]观察了12个牡丹品种的核型,分为3种类型;文献[18]比较了甘肃紫斑牡丹与中原牡丹品种染色体核型差异,其中,中原牡丹品种核型组成为2n=2x=6m+2sm+2st,甘肃紫斑牡丹品种核型组成为2n=2x=8m+2st。本文所研究的14个品种与文献[17]一致,亦分为3种类型。这说明:原产中国的牡丹存在着核型的多样性。但是,核型与牡丹株型、花型、花色等性状间无明显的相关性。
3.2 关于随体问题
本文研究表明:牡丹品种‘十八号’、‘银粉金鳞’的第5对染色体短臂上及‘红梅傲霜’第4对染色体短臂上发现有随体存在,其他品种未发现。这与文献[17]观察结果一致。但文献[16]对7个牡丹品种、文献[18]对10个牡丹品种及文献[19]对‘凤丹白’的观察并未发现随体存在。
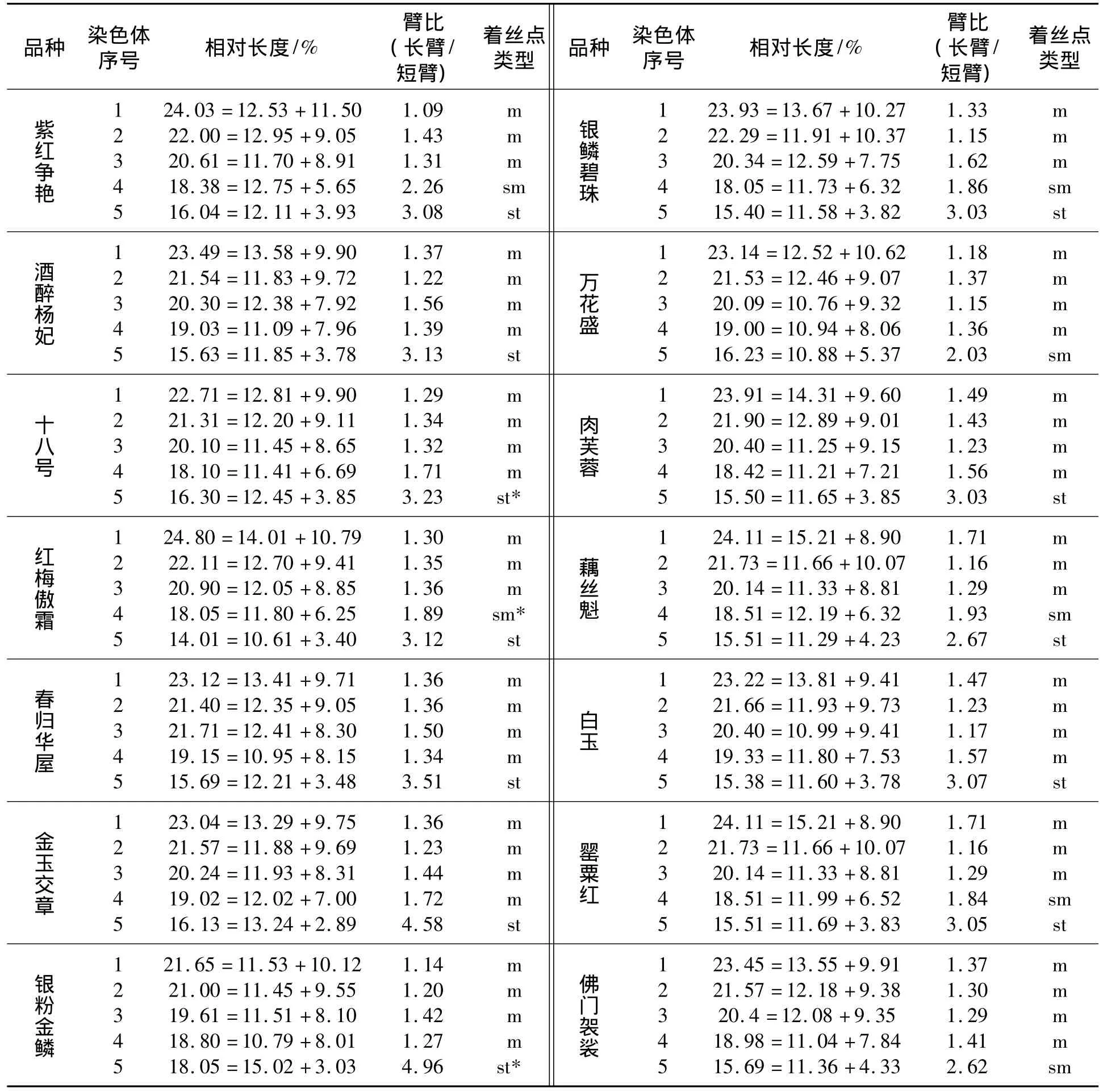
表2 牡丹品种核型特征

图1 牡丹品种核型图
4 结论
供试的14个品种间染色体相对长度差异较小,但核型组成不同,它们分属于3种核型类型。在3个品种中发现有随体存在。但这些差异与牡丹品种间株型、花型、花色等性状无明显的相关性。究竟牡丹株型、花型及花色等性状与染色体行为是否相关,有待于进一步研究。
[1] 洪德元,潘开玉.芍药属牡丹组的分类历史和分类处理[J].植物分类学报,1999,37:351-368.
[2] 李嘉珏,张西方,赵孝庆.中国牡丹[M].北京:中国大百科全书出版社,2011.
[3] 劳世辉,盛鸥,魏岳荣,等.香蕉A基因组6个品种的核型分析[J].园艺学报,2012,39(3):436-442.
[4] 张芳,邢世岩,韩晨静,等.叶籽银杏种质资源染色体核型分析及进化趋势[J].园艺学报2011,38(12):2245-2252.
[5] 孙晓梅,贾莲,杨宏光,等.新铁炮百合3个品种的核型分析[J].园艺学报,2010,37(6):1003-1008.
[6] 杨涤清,朱燮桴.草芍药、野牡丹和黄牡丹的核型研究[J].云南植物研究,1989,11(2):139-144.
[7] 李思锋,于兆英,周俊彦.黄牡丹的核型分析[J].武汉植物研究,1989,7(2):107-111.
[8] 肖调江,龚洵.滇牡丹复合群的GiernsaC-带比较研究[J].云南植物研究,1997,19(4):395-401.
[9] 裴颜龙.牡丹复合体的研究[D].北京:中国科学院植物研究所,1993:57.
[10] 张赞平,侯小改.杨山牡丹的核型分析[J].遗传,1996,18(5):3-6.
[11] 于兆英,李思锋,周俊彦.珍稀植物—紫斑牡丹和矮牡丹的核型分析[J].西北植物学报,1989,7(1):12-16.
[12] 洪德元,张志宽,朱相云.芍药属研究(1)—国产几个野生种核型报道[J].植物分类学报,1988,26(1):33-43.
[13] Zhu XW.The Karyotype of Five Cultivars in Paeonia Suffruticosa TronSino-Jpn Symposium TL[J].Chromos Research,1987:375-379.
[14] 龚洵,顾志建,武全安.黄牡丹七居群的细胞学研究[J].云南植物研究,1991,13(4):402-410.
[15] 龚洵,肖调江,顾志建.黄牡丹八个居群的Giemsa C-带比较研究[J].云南植物研究,1999,21(4):477-482.
[16] 王莲英,刘淑敏,秦魁杰,等.牡丹及其栽培品种的染色体组型[J].北京林学院学报,1983(1):63-69.
[17] 张赞平,张益民.栽培牡丹的染色体数目和核型变异[J].河南农业大学学报,1989,23(1):48-52.
[18] 于玲,何丽霞,李嘉珏.甘肃紫斑牡丹与中原牡丹类群染色体的比较研究[J].园艺学报,1997,24(1):79-83.
[19] 李子峰,王佳,胡永红,等.凤丹白牡丹核型分析与减数分裂的细胞遗传学观察[J].园艺学报,2007,34(2):411-416.
[20] 何丽霞,李睿,张延东.杨山牡丹×紫斑牡丹杂交后代“梦幻”的细胞学研究[J].兰州大学学报:自然科学版,2010,46(4):70-75.
[21] 李懋学,张赞平.作物染色体及其研究技术[M].北京:中国农业出版社,1996.
[22] Levan A,Fredga K,Sandberg A A.Nomenclature for Centromeric Position on Chromosomes Hereditas[J].Hereditas,1964,52(2):201-220.
[23] Stebbins G L.Chromosomal Evolution in Higher Plants[M].London:Edward Arnold,1971.
[24] Stebbins G L.Cytogenetie Rtudies in PaeoniaⅡ.The Cytoogg of the Diploid Species and Hybrids[J].Geneties,1938,23:53-110.
